
Биологический урожай это сумма приростов вещества в течение вегетационного периода
Обновлено: 05.10.2024
Органическое вещество, составляющее биомассу растений, образуется в процессе фотосинтеза из углекислого газа и воды за счет энергии поглощенной растениями ФАР. Связанная в урожае энергия ФАР является, таким образом, энергетическим выражением продуктивности фитоценоза.
Уже первые исследования показали, что между интенсивностью фотосинтеза единицы площади листа в единицу времени и накоплением биомассы растения отсутствует прямая зависимость (Тагеева, 1931; Катунский, 1941). В балансовом уравнении Иванова (1941) общая продуктивность фотосинтеза рассматривается как функция средней за учетный период интенсивности фотосинтеза величины фотосинтезирующей поверхности и рабочего времени. Уравнение учитывает также интенсивность дыхания и объем дышащей массы растения.
С развитием работ по проблеме "Фотосинтез и урожай" возникло представление о фотосинтетической деятельности растений как сложной совокупности процессов, в основе которых лежит поглощение растениями в фитоценозах энергии ФАР, и использование ее в процессе фотосинтеза на формирование урожаев (Ничипорович, 1956, 1966, 1966а, 1977).
Фотосинтетическая деятельность растений характеризуется интенсивностью фотосинтеза листьев и качественным составом образующихся в них биопродуктов, ходом роста вегетативных органов и листовой поверхности, накоплением биомассы растений, распределением продуктов фотосинтеза между вегетативными и репродуктивными органами и др.
Фотосинтетическая деятельность растений находится под постоянным влиянием состояния и динамики внешней среды. В отличие от одиночно стоящего растения на фотосинтетическую деятельность растений в ценозах оказывает влияние ценотическое взаимодействие растений, проявляющееся в конкуренции растений за условия жизни.
Как процесс формирования урожая фотосинтетическая деятельность растений подчиняется определенным законам и поддается количественному и качественному описанию, моделированию и управлению.
В основе работ по программированному получению урожаев лежит уравнение урожая, предложенное Ничипоровичем (1956). Уравнение дает количественное выражение зависимости биологических урожаев от размеров и работы листового аппарата с учетом качественной направленности процесса:
где Убиол — общий вес сухой биомассы растения; Фсо2 — интенсивность фотосинтеза (количество усвоенной СO2 в g/m2.день); Кэф— коэффициент эффективности фотосинтеза (отношение фактически накопленной сухой массы урожая к количеству усвоенной СO2); Л—площадь листьев (m2/ha); и — число дней продукционного периода.
Общий биологический урожай (Убиол) представляет собой сумму суточных приростов сухой массы растения. Зависимость хозяйственного урожая (зерно, плоды и др.) от биологического урожая представляется уравнением
где Кхоз — коэффициент хозяйственной эффективности фотосинтеза, характеризующий долю сухого вещества Ухоз от массы Убиол.
Для характеристики фотосинтетической деятельности растений наряду с размерами площади листьев используется показатель фотосинтетического потенциала (ФП). Он выражается в m2. днях и представляет собой сумму ежедневных площадей листьев растения или ценоза за вегетационный период или за какую-то его часть (ФП= ΣЛ.п). Показатель ФП является одним из основных при программировании урожая. Выражение ФСO2 х Kэф характеризует чистую продуктивность фотосинтеза (ЧПФ, или ,,нетто-ассимиляцию“), т. е. весовое количество суточного прироста сухого вещества растения в расчете на 1 m2 площади листьев. С учетом указанных преобразований уравнение (5) получит следующий вид:
Суточный прирост общей сухой массы биологического урожая определяется как произведение
На основе данных по фотосинтетической деятельности (Л, ФП, ЧПФ, С) с учетом факторов обеспечения (вода, минеральное питание и др.) определяется предполагаемое или планируемое нарастание биомассы по фазам вегетации на протяжении вегетационного периода для получения заданной величины Убиол с оптимальной структурой урожая (Кхоз).
Наивысшие урожаи могут быть получены при следующих оптимальных условиях (Ничипорович, 1956):
а) быстром развитии и сохранении в течение возможно длительного времени в активном состоянии большей площади листьев (Л);
б) наиболее высоких величинах интенсивности и коэффициентах эффективности фотосинтеза;
в) наиболее высокой чистой продуктивности фотосинтеза (ЧПФ) и высоких суточных приростах (С) сухого вещества;
г) наилучшем распределении и использовании образуемых веществ на формирование хозяйственной части урожая (Кхоз);
д) наиболее полном использовании потенциального периода вегетации (n).
Таблица 1
Зависимость величины урожая винограда (kg) от площади листьев (т2)
1 kg сахара ягод
Kozina (цит. по Стоеву, 1973)
Couvillon, Nakayama, 1970
Liuni (цит. по Стоеву, 1973)
Тодоров, Занков, 1964
Мельник, Анисимова, 1953
Для программирования урожая винограда представляет интерес показатель "продуктивности листьев", характеризующий количество сырого урожая или сахара гроздей 1 , произведенное в расчете на единицу площади листьев.
Следует отметить, что простой пересчет сырой массы или валового сахара гроздей на листовую площадь растения характеризует фактически сложившееся соотношение между площадью листьев и гроздями, но не потенциальную продуктивность листьев (ПЛ). Для получения характеристики потенциальной ПЛ необходимо, чтобы площадь листьев по отношению к гроздям находилась в оптимуме или ограничивала бы рост гроздей. Это достигается в опытах с различной нагрузкой побегами или путем частичного удаления листьев с побега при одновременном их кольцевании (Амирджанов, 1963).
В табл. 1 показаны полученные разными исследователями данные по продуктивности листьев винограда. Для возможности сопоставления они приведены нами к одной характеристике: площади листьев, необходимой для получения единицы урожая (1 kg сырой массы гроздей или 1 kg сахара гроздей).
Величина ПЛ винограда варьирует в широких пределах; от 3 до 15 m2/kg сахара и от 0,7 до 2,0 m2 /kg сырого урожая гроздей, что, очевидно, в большей степени вызвано разными методами расчета, чем фактической потенциальной продуктивностью листьев. Если с известным допущением усреднить соответствующие крайние значения, то необходимая площадь листьев составит для получения 1 kg сырого урожая ягод — 1—1,5 m2; для получения 1 kg сахара гроздей — 6—9 m2.
Эти величины практически идентичны для ягод с сахаристостью сока 17—18% при содержании сока в ягодах 80%.
Таким образом, по самой приближенной оценке при благоприятных условиях для получения 100 cwt гроздей при сахаристости сока ягод 17—18 % необходимо иметь на гектаре - 10—15 тыс. m2 площади листьев. Количество побегов, соответствующее этой площади листьев и необходимое для получения заданного урожая, будет зависеть от фитометрических характеристик побега и показателей его плодоносности: коэффициента плодоношения 2 и средней массы грозди или, иначе говоря, от продуктивности одного побега.
Как показывают исследования, урожаи наиболее тесно коррелируют с размерами листовой поверхности. Следовательно, для получения планируемого урожая винограда необходимо иметь на гектаре такое количество нормально развитых побегов, ход роста которых обеспечил бы получение расчетной величины ФГТ. Это обстоятельство указывает на важность изучения закономерностей роста побегов винограда в зависимости от нагрузки побегами и гроздями, длины обрезки, формировки, фона питания и др. с целью установления "оптимального" хода роста.
При оптимальных условиях произрастания растения должны не только осуществлять большую фотосинтетическую работу, но и эффективно использовать продукты фотосинтеза на формирование хозяйственной части урожая— гроздей.
У винограда структура биологического урожая определяется как внешними факторами, так и природой самого растения. К числу "внутренних" факторов относятся такие, как сила роста куста, плодоносность побегов, величина грозди, нагрузка побегами и гроздями и др. У растений с одинаковой вегетативной массой величина Кхоз будет тем больше, чем выше плодоносность побегов. При одинаковой плодоносности уменьшение вегетативного прироста повлечет за собой повышение показателя Кхоз.
Величина Кхоз у винограда изменяется в больших пределах. У сорта Алиготе в зависимости от площади питания она изменялась от 0,45 до 0,57; у сорта Карабурну — от 0,60 до 0,67 (Плакида, 1967). У сорта Болгар (синоним Карабурну) в зависимости от высоты штамба величина Кхоз, изменялась от 0,48 до 0,71 (Панделиев, 1976). В опытах Стоева и Добревой (1976) у сорта Ркацители в приземной культуре Кхоз составил 0,45, а при штамбовой культуре — 0,73 при близких значениях Убиол (1,34 и 1,38 kg на куст).
1 Поскольку процентное содержание сахара в соке ягод в разных случаях неодинаково, правильнее характеризовать этот показатель не по сырому урожаю, а по количеству сахара гроздей.
2 Коэффициент плодоношения (Кпл) — среднее число гроздей на один развившийся побег.
По нашим данным у сорта Шабаш в зависимости от густоты посадки и формировки кустов величина Кхоз изменялась от 0,33 до 0,46; у сорта Рислинг рейнский — от 0,33 до 0,52. У сортов Пино гри, Мускат белый и Хиндогны в среднем за пять лет величина Кхоз составила соответственно 0,30, 0,47 и 0,36.
Задача состоит в том, чтобы при помощи различных агротехнических приемов находить для каждого отдельного случая оптимальную величину Кхоз, т. е. добиваться наилучшего перераспределения ассимилятов на формирование хозяйственного урожая. Вместе с тем величину Кхоз у винограда нельзя увеличивать беспредельно. Превышение Кхоз некоторого оптимального для конкретного случая уровня практически будет означать перегрузку кустов гроздями, и это может служить причиной ухудшения качества ягод и ослабления силы кустов. С точки зрения создания высокопродуктивных насаждений с оптимальной структурой кустов важным представляется подбор сортов, сочетающих высокие показатели плодоносности с относительно умеренным ростом побегов.
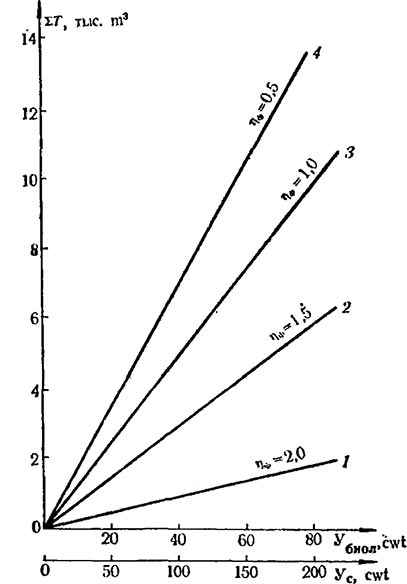
Получение планируемых урожаев винограда требует знания механизма взаимодействия показателей фотосинтетической деятельности. Пример такого взаимодействия показан на рис. 1.
Рис. 1. Графическое представление механизма фотосинтетической деятельности винограда в насаждении
1 — сухая биомасса (У); 2 — потенциальный хозяйственный урожай (Ухоз); 3 — хозяйственный коэффициент (Кхоз); 4 — чистая продуктивность фотосинтеза (ЧПФ); 5 — реальный хозяйственный урожай (Ухоз 2)
С увеличением на гектаре площади листьев (ФП) растет величина Убиол, но не пропорционально, поскольку с ростом площади листьев снижается ЧПФ. По мере роста площади листьев (Л) повышается также величина Ухоз. Максимальные уровни Ухоз при заданных Л характеризуются линией регрессии Ухоз 1, которая получена при условии, что величина Кхоз оставалась постоянной (0,45 для сорта Шабаш). Однако по мере роста Л снижается Kхоз, и в результате реальные уровни хозяйственного урожая — Ухоз 2 отклоняются от максимальных. По мере увеличения Л насаждения разрыв между максимальными (Ухоз 1) и реальными (Ухоз 2) урожаями гроздей увеличивается; возникает как бы „эффект ножниц“. Уменьшение Кхоз в насаждениях с густой посадкой (имеющих высокие показатели Л) обусловлено снижением плодоносности побегов: Кпл и средней массы грозди.
Для разбираемого случая зависимость Убиол (cwt/ha) от ФП (млн.m2. дней) имеет линейный характер и описывается уравнением
Связь между Ухоз (cwt сухого вещества) и площадью листьев (тыс. m2) описывается квадратичным уравнением
Аналогичный характер связи выявлен Ординовой (1968) между сырым урожаем гроздей и густотой посадки виноградника. Следовательно, одной из задач оптимизации продукционного процесса винограда для получения планируемых урожаев является отыскание оптимальных размеров площади листьев насаждения (количества побегов и соответствующего их размещения в пространстве), обеспечивающих получение максимального хозяйственного урожая без снижения его качества. Планируемый урожай должен быть обеспечен водой и минеральным питанием.

В процессе фотосинтеза в растении образуются и накапливаются органические вещества. Количество их зависит от интенсивности фотосинтеза и дыхания растений, от соотношения между этими процессами, от положения так называемой компенсационной точки. Это такое состояние, когда в растении фотосинтез и дыхание полностью уравновешиваются, оно означает, что сколько вещества создано, столько и израсходовано в процессе дыхания, т. е. нет ни увеличения, ни уменьшения массы, окружающая атмосфера не обогащается ни кислородом, ни углекислым газом. При таких условиях органические вещества не накапливаются.
Накопление органического вещества растением за определенный период времени или за всю его жизнь следует рассматривать как разницу между количеством созданного на свету органического вещества и израсходованного на дыхание. Кроме того, во время вегетации в растении могут происходить превращения продуктов фотосинтеза с изменением их массы. Например, масса откладываемой в запас клетчатки на 10% меньше, чем масса глюкозы, из которой она образуется. К этому следует добавить, что отмирание корневых волосков и мелких корешков, сбрасывание части цветков — потери, происходящие в период вегетации. Поэтому общая продуктивность растения будет определяться не только образованием органических веществ в процессе фотосинтеза, но и величиной всех указанных потерь.
Доказано, что энергия, используемая для образования продуктов фотосинтеза при благоприятных условиях (высокой концентрации СО2 в окружающем воздухе, хорошем водоснабжении и т. д.), составляет всего 2-5%. Обычно при высокой интенсивности радиации количество энергии, затраченной на фотосинтез, равно 1%, или 16 кДж/м 2 в 1 ч.
На основании ряда исследований, в частности работ английских ученых Броуна, Эскомба и Вильсона, можно утверждать, что при нагревании листа растение может отдавать в виде теплоизлучения 84 кДж/м 2 в 1 ч на каждый градус температурного градиента между листом растения и окружающим воздухом. Остальная энергия в количестве 1500 кДж/м 2 в 1 ч (99% энергии, или 1584 кДж без энергии теплоизлучения), превращаясь в тепло, способствует нагреванию и испарению воды из листьев — транспирации, требующей большого расхода энергии. На испарение 1 г воды при 20°С затрачивается 2,45 кДж (скрытая теплота испарения воды). Растение, поглощая 1500 кДж/м 2 в 1 ч, может испарить 612 г воды, что соответствует транспирационному коэффициенту ряда растений.
Подсчеты коэффициента использования солнечной энергии показали, что максимальный прирост сухого вещества на одно растение кукурузы в период выбрасывания метелки за сутки составляет 20 г. Принимая это сухое вещество за крахмал, что почти соответствует действительности, и зная, что теплота сгорания 1 г крахмала составляет 16,6 кДж, можно рассчитать суточный прирост энергии на одно растение кукурузы, который будет составлять 351 кДж. На 1 га при площади питания 70×70 см размещается 30-40 тыс. растений кукурузы. Таким образом, за сутки накопится 10534×10 3 —14045×10 3 кДж энергии. За 6 ч на поверхность 1 га посевов кукурузы в летний период поступает 209 млн кДж, из них растениями кукурузы используется лишь 5-6% энергии солнечной радиации. Расчеты показали, что на создание урожая яровая пшеница использует 3,26%, картофель — 3,02, сахарная свекла — 2,12% лучистой энергии.
Количество ФАР, поглощенной посевом, растительным покровом, определяют по формуле, Дж/см 2 в 1 мин: $$\large \Pi = Q - R - T_n + R_n$$
где П — поглощенная посевом радиация; Q — суммарная радиация, падающая на посев; R — радиация, oтраженная от посева и вышедшая за пределы его верхней границы; Тn — радиация, проникшая к почве; Rn — радиация, отраженная от почвы под растительностью.
Коэффициент поглощения энергии ФАР (Qп) посевом определяют делением обеих частей формулы на Q: $$\large Q_\pi = \frac<\Pi> = 1 -\frac - \frac + \frac, $$
где \(\frac\) — альбедо посева, показывающее, какая доля падающей радиации у отражается посевом;\(\frac\) — коэффициент пропускания, показывающий, какая доля падающей радиации (Q) достигает почвы под растительностью; \(\frac\) — альбедо почвы под растительностью (К. Г. Тооминг, Б. Н. Гуляев).
Эффективность фотосинтеза можно характеризовать коэффициентом полезного действия (КПД), который определяют по формуле: $$\large E \% (КПД) = \frac,$$
где А — количество энергии, поступившей за период вегетации па 1 га посева, или энергии, которая была поглощена посевом, кДж; В — количество энергии, накопившейся в органической массе урожая (биологического или хозяйственного), кДж.
В среднем КПД фотосинтеза сельскохозяйственных растений составляет 0,5-1%, а теоретически возможный — 4-6%. Максимальный показатель использования энергии ФАР посевами и насаждениями обусловливается способностью поглощать не менее 60% энергии света, поступающего к ним на протяжении вегетации. Из этого количества на фотосинтез идет 10%, на дыхание расходуется около 20% энергии, усваивающейся в процессе фотосинтеза, или $$\large E \% (КПД) = 60 \cdot \frac \cdot \frac = 48 \%.$$
В 1 кг сухой массы урожая накапливается до 17 тыс. кДж энергии.
Поступающая солнечная энергия в разных зонах неодинаково взаимодействует с режимом температуры и влаги, вследствие чего значительно изменяется эффективность солнечной радиации как фактора фотосинтеза и продуктивности. Кроме того, следует отметить, что она является не только движущей силой фотосинтеза, но и основным фактором транспирации. Свет влияет и па такие физиологические процессы, как рост, органогенез, передвижение ассимилятов. Один из основных путей повышения продуктивности фотосинтеза — увеличение до определенных размеров площади ассимилирующих органов — листьев и усиление их деятельности, что обусловливает количество лучистой энергии, поглощаемой хлоропластами. Доказано, что величина урожая в значительной мере зависит от оптималык структуры посевов. Структурой посевов называется создаваемая архитектоника (построение) сообщества растений, которое характеризуется определенными морфологическими признаками и физиологическими функциями, а оптимальная структура — это такой посев, который имеет высокий КПД фотосинтеза и обеспечивает максимальный биологический и хозяйственный урожай.
| Вариант | Дни после всходов | ||||||||
| 27 | 36 | 46 | 59 | 68 | 87 | ||||
| Площадь листьев, тыс. м 2 | |||||||||
| Без подкормки | 6,6 | 11,4 | 17,2 | 19.8 | 19,8 | 19,3 | |||
| Подкормлено NPK | 7,4 | 11,8 | 20,3 | 21.9 | 21,3 | 19,6 | |||
| Поглощено ФАР, тыс. кДж/мин | |||||||||
| Без подкормки | 11,0 | 84,9 | 109,3 | 130,8 | 141,7 | 137,5 | |||
| Подкормлено NPK | 68,1 | 91,1 | 139,6 | 157,2 | 163,9 | 149,2 | |||
Установлено, что при подкормке растений увеличиваются размеры листовой поверхности, а также изменяются физиологические особенности фотосинтетического аппарата— способность поглощать и усваивать лучистую энергию. Так, более развитая листовая поверхность подкормленных растений кукурузы в период вегетации поглощала и большее количество ФАР (табл. 10). Кроме того, листья кукурузы разных ярусов неодинаково поглощают лучистую энергию. Распределение ФАР внутри посева непропорционально площади листьев по отдельным ярусам. Например, верхний слой листьев составляет лишь 23,7% всей листовой поверхности, а поглощает 47% энергии, в то время как средний слой, составляющий 60,4% этой поверхности, усваивает только 36,6% поглощенной посевом ФАР. В результате подкормки растений NPK уменьшилось количество энергии, поглощаемой нижним ярусом листьев, с 26,4 до 8,6% и увеличился процент поглощения ФАР средним слоем — с 36,6 до 49,7%. Таким образом, условия минерального питания в значительной мере влияют на структуру посевов и поглощение ФАР листьями разных ярусов (слоев травостоя).
Повышение фотосинтетической продуктивности растений при бесперебойном снабжении их водой и питательными веществами обеспечивается наиболее целесообразным размещением растений на площади, т. е. так, чтобы они в достаточной степени освещались солнечными лучами. Поэтому в сельскохозяйственной практике используют различные способы посева. Для лучшего освещения растений, а также для более эффективного использования лучистой энергии рядки располагают с востока на запад или с северо-востока на юго-запад.
Таким образом, в течение вегетации необходимо контролировать и регулировать интенсивность фотосинтеза, энергетический баланс посева, КПД фотосинтеза за короткие промежутки времени, прирост общей сухой массы. В связи с этим поставлена задача создавать и внедрять в производство сорта, способные развивать большую листовую поверхность, имеющие высокую продуктивность фотосинтеза и дающие большой биологический и хозяйственный урожай.
Предположим, что интенсивность фотосинтеза будет не 10-15 мг, а 30-50 мг СО2 на 1 дм 2 в 1 ч, тогда при средней площади листьев 30-40 тыс. м 2 на 1 га у таких растений, как сахарная свекла, картофель, урожай корне- и клубнеплодов, будет составлять 6-8 т с 1 га (в пересчете на сухую массу).
Абсолютная величина продуктивности растения равна в среднем 40-50 мг органического вещества в сутки на 50 см 2 листовой поверхности при условии, что дыхание составляет не более 5-10% фотосинтеза. Фотосинтез в естественных условиях — процесс очень изменчивый, и по основным его показателям — интенсивности (мг СО2 на 1 дм 2 в 1 ч) и чистой продуктивности (г/м 2 в сутки) — наблюдается большая вариабельность. Чистую продуктивность фотосинтеза (Фч.пр), которая характеризует интенсивность образования и накопления органической массы урожая, определяют по формуле, г/м 2 : $$\large Ф_ = \frac<(Л_1 + Л_2) \frac<1>n> ,$$
Если, кроме общей чистой продуктивности фотосинтеза, надо определить интенсивность работы листьев и ее направленность на формирование хозяйственно ценных органов, например корнеплодов, клубней или зерна, в уравнение вместо В2 — B1 подставляют величину Х2 — X1, т. е. прирост сухой массы хозяйственно ценной части урожая.
Фотосинтез посевов (Фп) можно оценить, используя значение суточных приростов сухой фитомассы по формуле, предложенной Б. И. Гулиевым и А. С. Оканенко: $$\large Ф_n = \frac,$$
Например, для посева кукурузы получены такие значения (мг С02 на 1 дм 2 в 1 ч): в Киеве — 54, в Волгограде — 84, в Москве — 18.
Потенциальную фотосинтетическую мощность посева может характеризовать фотосинтетический потенциал растений — сумма ежедневных показателей площади листьев посева за весь вегетационный период или часть его, выраженная в м 2 дней/га.
Различают урожай биологический и хозяйственный.
Биологический урожай — это сумма суточных приростов за весь вегетационный период. Его можно выразить формулой $$\large У_ = \Sigma C_,n.$$
где С — суточные приросты массы, кг/га в сутки. $$\large C = \fracЛ>,$$
где Ф — количество ассимилированного СО2; Кэф — коэффициент фотосинтеза $$\large \left ( \frac>\; CO_2> \right);$$
Л — листовая поверхность; 1000 — граммы.
Хозяйственный урожай составляет определенную долю биологического. Не все части растения равноценны и одинаково используются. Например, у хлебных злаков наиболее ценным является зерно, у сахарной свеклы — корень и т. д. Таким образом, у различных культур коэффициент хозяйственного использования (Кхоз) разный. Поэтому хозяйственный урожай равен: $$\large У_ = У_К_.$$
Таким образом, дальнейшее повышение урожайности сельскохозяйственных культур должно осуществляться путем увеличения продуктивной части растения. Знание закономерностей, определяющих те или иные изменения интенсивности и продуктивности фотосинтеза, умение управлять этими изменениями — одна из важных основ получения больших биологических и хозяйственных урожаев высокого качества.
Продуктивность фотосинтеза сельскохозяйственных культур целесообразно оценивать величиной выхода полезной энергии с одного гектара.
Полезная энергия — это материализованная солнечная энергия в урожае за вычетом технологической энергии, затраченной на его производство (обработка почвы, удобрение, сорт, приемы агротехники).
При интенсивной технологии возделывания сельскохозяйственных культур, предусматривающей высокий КПД фотосинтеза и высокую урожайность, минимальные затраты технологической энергии, количественное соотношение содержащейся в урожае и технологической энергии может служить важным показателем энергетической эффективности культуры, т. е. энергетических затрат на производство единицы сельскохозяйственной продукции.
Фотосинтез растений изучается очень активно. В нашей стране ведутся широкие исследования по физико-химическим и прикладным проблемам этого процесса.
Познание природы фотосинтеза растений — одна из наиболее важных, фундаментальных проблем биологии, тесно связанных с удовлетворением нужд человечества в пище и энергии.
Значительные результаты достигнуты в изучении молекулярных механизмов преобразования энергии света при фотосинтезе. Установлено, что этот процесс начинается с поглощения энергии света (фотонов) пигментами антенны и миграции энергии возбуждения к реакционным центрам, где за пикосекунды происходит преобразование ее в химическую энергию; изучаются улътрамолекулярная структура реакционного центра и функции его компонентов; решается задача построения искусственных реакционных центров; большое внимание уделяется изучению механизма переноса электронов в фотосинтезирующих системах.
Ведутся исследования по направленному синтезу продуктов фотосинтетического метаболизма (углеводов, белков, жиров). Большое внимание уделяется применению принципов фотосинтеза для построения искусственных систем, моделей, использующих энергию света. Важное значение придается созданию биотехнических систем, потребляющих солнечную энергию для образования водорода и восстановления молекулярного азота, а также созданию фотобиотехнических систем, использующих солнечную энергию с предельно достижимым КПД (до 30%). Исследуются молекулярные фотокаталитические системы как преобразователи солнечной энергии при высоких концентрациях СО2 и высоких интенсивностях света.
По комплексной программе ведутся исследования бактериального фотосинтеза в направлении молекулярной организации фотосинтетических систем, поглощения, переноса и конверсии энергии света.
Выполнение всех этих и ряда других фундаментальных исследований внесет значительный вклад в теорию и практику фотосинтеза.
Документ из архива "Физиология растений", который расположен в категории "учебные пособия". Всё это находится в предмете "биология" из раздела "Книги, пособия, лекции и семинары", которые можно найти в файловом архиве Студент. Не смотря на прямую связь этого архива с Студент, его также можно найти и в других разделах. Архив можно найти в разделе "книги и методические указания", в предмете "биология" в общих файлах.
Онлайн просмотр документа "10802"
Текст 9 страницы из документа "10802"
Основываясь на механизмах влияния внутренних и внешних факторов, действующих на показатели фотосинтетической активности растений, в практике сельского хозяйства используют ряд приемов, позволяющих увеличить интенсивность фотосинтеза и повысить урожайность сельскохозяйственных культур.
Прежде всего это точное соблюдение оптимальной технологии:
соблюдение режима орошения,
соблюдение режима минерального питания,
использование необходимых внекорневых подкормок микроэлементами,
повышение в защищенном грунте концентрации углекислого газа за счет применения органических удобрений (внесение навоза), использования сухого льда, поддымление парниковых рам. При этом у огурцов не только повышается интенсивность фотосинтеза, но и увеличивается количество женских цветков.
Соотношение между количеством усвоенного в процессе фотосинтеза углекислого газа и накопленного сухого органического вещества называется коэффициентом эффективности фотосинтеза.
Необходимо учитывать, что на итоговое накопление органического вещества влияют два процесса: фотосинтез и дыхание. Количество накапливаемых органических веществ зависит от интенсивности фотосинтеза и дыхания растений, то есть от положения компенсационной точки. Компенсационная точка характеризует такое состояние растения, когда в нем фотосинтез и дыхание полностью уравновешиваются, т.е. при таких условиях органическое вещество не накапливается.
Накопление органического вещества растением за определенный период или за всю его жизнь следует рассматривать как разницу между количеством созданного на свету органического вещества и израсходованного на дыхание.
Кроме того, в процессе преобразования веществ также может происходить уменьшение массы (например, масса клетчатки на 10% меньше массы глюкозы, из которой она образуется), на накопление органического вещества влияет также опадение или гибель частей растения (цветков, корневых волосков) в период вегетации.
При оптимальных условиях влажности и температурного режима важно также соблюдать оптимальное размещение растений. Для лучшего освещения растений рядки располагают с востока на запад или с северо-востока на юго-запад. Величина урожая в значительной мере зависит от оптимальной структуры посевов.
Структурой посевов называется создаваемая архитектоника сообщества растений, которое характеризуется определенными морфологическими признаками и физиологическими функциями, а оптимальная структура - это такой посев, который имеет высокий КПД фотосинтеза и обеспечивает максимальный урожай. Обычно потери энергии на дыхание составляют 15-25%, но при загущенном посеве нижние, а частью и средние листья становятся не столько синтезирующими, сколько потреблояющими.
Одной из важнейших задач селекции является создание сортов, способных развивать большую фотосинтезирующую поверхность, имеющих высокую продуктивность фотосинтеза и дающих большой биологических и хозяйственный урожай.
В сельском хозяйстве наибольший интерес представляет получение конечного продукта - полезной накопленной биомассы растений, то есть чистой продуктивности фотосинтеза.
Чистую продуктивность фотосинтеза определяют, пользуясь легко определяемыми величинами: площадью листовой поверхности и фактически накопленной биомассой:
где В1 и В2 - масса сухого вещества пробы урожая в начале и конце учетного периода, т.е.
В2 - В1 - прирост сухой массы за учетный период (п дней), Л1 и Л2 - площадь листьев пробы в начале и конце периода, т.е. (Л1+Л2).1/2п - средняя площадь листьев за указанный отрезок времени, п - число дней в учетном периоде.
Кроме общей чистой продуктивности фотосинтеза определяют и интенсивность работы листьев, направленную на создание хозяйственной части урожая. В этом случае вместо В2 - В1 подставляют величины Х2 - Х1, то есть прирост сухой массы хозяйственной части урожая.
Фотосинтетический потенциал растений - это сумма ежедневных показателей площади листьев посева за весь вегетационный период (или за его часть), выраженная в
м 2. дни/га.
Биологический урожай - это сумма суточных приростов за весь вегетационный период:
Убиол. = С1,2. п,
где С - суточные приросты массы, в кг/га в сутки.
Ф . Кэф.
Х1000,где Ф - количество ассимилированного СО2, Кэф. - коэффициент фотосинтеза (суточный прирост урожая/ суточное усвоение углекислого газа), Л - листовая поверхность, 1000 - граммы (для пересчета в кг). В среднем при хорошей агротехнике сельскохозяйственные культуры накапливают 15-20 т/га сухой биомассы.
Хозяйственный урожай составляет ту долю биологического, которая используется человеком:
Ухоз. = Убиол. Кхоз.
Коэффициент хозяйственного использования у разных культур может сильно различаться (зерновые культуры и сахарная свекла).
Продуктивность фотосинтеза сельскохозяйственных культур целесообразно оценивать величиной выхода полезной энергии с гектара.
Демонстрация таблиц с данными по интенсивности фотосинтеза для разных культур.
Для определения площади листовой поверхности используют три основных метода:
фотопланиметрирование - когда с помощью специального прибора определяют уменьшение интенсивности светового потока, пропорциональное площади листа,
расчет по высечкам - когда, взвесив несколько высечек известной площади, делят общий вес листа на вес высечек и узнают общую площадь листа,
расчет по линейным размерам листа по формуле S=а . b . k, где а и b - ширина и длина листа, а k - коэффициент для данной сельскохозяйственной культуры. Для всех сельскохозяйственных культур он определен и приведен в справочниках: для кукурузы - 0,68; для ячменя - 0,65; для сахарной свеклы - 0,76; для яблони - 0,62-0,74.
Суточный ход фотосинтеза у светолюбивых и теневыносливых растений.
В связи с тем, что основой жизнедеятельности растений является фотосинтез, все растения любят свет. Однако, есть растения, выносливые к недостатку света, которые называют теневыносливыми. Светолюбивость или теневыносливость растений определяется сформировавшимися в процессе эволюции механизмами приспособления к окружающим факторам среды.
Светолюбивость проявляется в значительном уменьшении содержания зеленых пигментов и соответствующем повышении количества каротиноидов. У светолюбивых растений часто наблюдается усиленное развитие ассимилирующей ткани палисадной паренхимы, состоящей из нескольких слоев клеток меньшей величины, чем у теневыносливых; у них число устьиц больше и поэтому углекислый газ быстрее проникает внутрь листа. Внешне у светолюбивых растений листья всегда более мелкие и имеют более бледную зеленую окраску, чем теневыносливые.
Интенсивность фотосинтеза у светолюбивых растений повышается к полудню и уменьшается утром и вечером, у теневыносливых максимум фотосинтеза приходится на утро и вечер, и минимальна интенсивность фотосинтеза в полдень.
Роль зеленых растений в природе.
Зеленые растения, являясь основными автотрофами на нашей планете, определяют несколько существеннейших параметров жизни:
накопление кислорода в атмосфере,
основное звено синтеза органики в пищевых цепях,
преобразование физической энергии Солнца в химическую энергию органических веществ.
В.И. Вернадский писал о значении растений так: "На нашей планете свободный кислород, находящийся на ней в виде газа или в форме раствора в природных водах, нацело создается жизнью. Мы не знаем пока ни одного случая, когда бы он выделялся в значительном количестве в результате какого бы то ни было химического процесса, независимо от жизни. Но нам известны тысячи земных химических процессов, в которых свободный кислород поглощается, переводится в новые соединения, исчезает как таковой. А между тем, количество его в биосфере не меняется, остается все тем же. Это достигается непрерывной работой зеленых растений, и это одно дает меру их значения на нашей планете, дает представление о размахе их геохимической энергии".
В ранние геологические периоды теплый и влажный климат и высокое содержание углекислого газа в атмосфере Земли содействовали пышному расцвету фотосинтезирующих растений. При этом все большее количество углерода переходило в состав органических веществ, которые затем превращались в залежи полезных ископаемых (уголь, нефть, торф) и гумус. Эти запасы ныне достигают 6 .1 0 15 тонн.
Космическая роль зеленых растений выражается в их влиянии на изменение условий жизни на Земле и в обеспечении возникновения новых форм жизни.
Атмосфера обогащалась кислородом, уменьшалось количество углекислого газа. Имеющееся в современной атмосфере количество кислорода, равное 1,5 .1 0 15 тонн, близко к тому количеству, которое должно было выделиться при образовании органических запасов углерода на Земле (6 .1 0 15 тонн).
По современным подсчетам годовая продуктивность всей растительности Земли оценивается в 100 миллиардов тонн сухой биомассы.
Преобладающую роль в продуктивности наземной растительности играют леса - 28,4 миллиарда тонн, затем идут пастбища (степи, луга) - 10,4 миллиарда тонн, затем сельскохозяйственные угодья, которые дают около 8,7 миллиардов тонн сухой биомассы.
Основное же количество сухой биомассы накапливает растительность океанов и других водоемов.
Ежегодно растения поглощают из окружающей среды 2.10 9 т азота, 6 .1 0 9 фосфора, 158 .1 0 9 т углекислого газа и 128 .1 0 9 т воды, а выделяют 115 .1 0 9 т кислорода.
В процессе дыхания растений окисляется 25 .1 0 9 т органики, причем выделяется 34,5 .1 0 9 т углекислого газа.
Весь животный мир разлагает 3,5 .1 0 9 т органики с образованием 4,8 .1 0 9 т углекислого газа.
За счет сжигания полезных ископаемых в промышленности и быту в атмосферу выделяется 15 .1 0 9 т углекислого газа.
Тема: Дыхание растений.
Дополнительная литература по теме:
В.Л. Кретович "Основы биохимии растений". М, 1971,А. Ленинджер "Биохимия" М., 1974,Я. Мусил, О. Новакова, К. Кунц "Современная биохимия в схемах".М., 1981.
Общая характеристика дыхания.
Цепь дыхательных ферментов.
Строение и функции митохондрии и дыхательных ферментов. Энергетика процесса дыхания.
Влияние условий окружающей среды на дыхание растений.
Регулирование дыхания сельскохозяйственных продуктов при хранении.
Общая характеристика дыхания.
Дыхание занимает исключительное положение среди других физиологических процессов. Окислительное дыхание свойственно всем многоклеточным живым организмам, как растительным, так и животным. Ряд видов прокариот также ведут этот процесс. Поэтому основные этапы дыхания являются одинаковыми для всех живых организмов, получающих энергию с помощью этого способа.
Дыхание является ключевым процессом метаболизма любого организма по двум причинам: при дыхании происходит освобождение химической энергии органических веществ, используемых в качестве дыхательного материала. Экзотермические реакции дыхательного процесса непосредственно связаны с эндотермическими процессами клеточного обмена и служат для них источником энергии. Таким образом, дыхание обеспечивает возможность течения эндотермических реакций обмена, процессов образования структур и осуществления движений, что требует затрат энергии, при дыхании протекают такие химические превращения, в результате которых образуются высокоактивные соединения, обладающие большой реактивной способностью и играющие исключительную роль в обмене веществ в организме.

Исследованиями А.А. Ничипоровича (1966) и А.Я. Бакалдина (1973) установлено, что урожай сухого вещества сельскохозяйственных культур на 80–90 % создается в результате фотосинтеза, который, в первую очередь, зависит от размеров ассимиляционной поверхности, высоты и густоты стояния растений и ряда других факторов. Все остальные процессы питания растений, в частности водное и минеральное, эффективны в той степени, когда они обеспечивают и поддерживают оптимальную деятельность фотосинтетического аппарата.
Нарастание сухой массы считается одним из главных показателей фотосинтетической деятельности растений. Суточный прирост сухого вещества значительно изменяется и может достигать до 300 кг/га в период интенсивного роста растений (А.А. Ничипорович и др., 1961).
Потенциальная биологическая продуктивность полевых культур зависит от величины поступающей к ним солнечной энергии и от особенностей её расхода. Помимо продуцирования кислорода, фотосинтез приобретает всё большее значение как основа воспроизводства продовольствия. Поэтому, создавая искусственные ценозы на полях, человек при помощи агротехники и лучистой энергии солнца обеспечивает себя продуктами питания. Первичные продукты фотосинтеза, участвуя в реакции вторичного метаболизма, образуют весь спектр органических соединений растительной клетки (А.Т. Мокроносов, В.Ф. Гавриленко, Т.В. Жигалова, 2006).
Высокая продуктивность сельскохозяйственных культур определяется динамически оптимальным соотношением отдельных элементов фотосинтеза. К основным из них относят: размер ассимиляционного аппарата, фотосинтетический потенциал, интенсивность и продуктивность фотосинтеза (Т.М. Русакова, 1974; J.M. Anderson, 1983; В.М. Важов, 2012).
Известно, что годовая масса органического вещества, создаваемого фотосинтезом, с избытком перекрывает то количество продовольствия, которое необходимо населению планеты. Поэтому одним из путей повышения эффективности земледелия является создание посевов с хорошей структурой пространственного расположения листьев.
Листовая поверхность является главным рабочим органом зелёных растений, а размер урожая почти полностью определяется её величиной. Нарастание листовой поверхности и величина ассимиляционного аппарата культурных растений зависит от агротехнических условий (А.Н. Бегишев,1953; Vong Hguyen Quoc, Murata Yoshio, 1978) .
По вопросу об оптимальной площади листовой поверхности сельскохозяйственных культур нет единого мнения. А.А. Ничипорович (1959) считает, что листовая поверхность должна составлять до 50 тыс. м2/га, а по мнению А.Н. Бегишева (1953) этот показатель равен примерно 100 тыс. м2/га.
В исследованиях А.Д. Гончарова (2008) в Новосибирской области бороздковые посевы формировали лучшую листовую поверхность гречихи, чем сплошные. С возрастом растений преимущество бороздковых посевов было очевидным. В фазу бутонизации площадь листьев на 1 га бороздковых посевов была больше рядовых на 14,5 %, а в фазу цветения – на 37,4 %. В изменении площади листьев по годам исследований каких-либо закономерностей не отмечено как по каждому способу возделывания, так и по сравнению их между собой. В различиях между изучаемыми способами посева по другим показателям фотосинтетической деятельности гречихи и их динамики в течение вегетационного периода в опытах А.Д. Гончарова (2008) проявились те же закономерности, что и в случае с площадью листьев. Так, фотосинтетический потенциал бороздковых посевов был больше соответствующего показателя сплошных посевов на 15 % за период бутонизация-цветение и на 30 % – за время цветение-созревание.
По мнению Н.Д. Кумсковой (2004), в условиях Дальнего Востока на листовую поверхность гречихи влияют сроки сева. В её опытах посевы 5 и 30 июня имели площадь листьев 22,5 и 23,5 тыс. м2 на 1 га, тогда как посевы 20 и 25 июня, соответственно, 32 и 31 тыс. м2. Гречиха давала хорошую вегетативную массу и высокие показатели фотосинтеза. В опыте Н.Д. Кумсковой (2004) при посеве с 5 по 30 июня получено от 25,1 до 37,3 т/га зелёной массы, максимальной она была при сроке сева 20 июня (табл. 56).
Фотосинтетическая деятельность и урожайность вегетативной массы гречихи
на разных сроках сева (по Н.Д. Кумсковой, 2004)
Площадь листьев (тыс. м2/га)
Зеленая масса, т/га
Сухая масса, т/га
Примечание. ЧПФ – чистая продуктивность фотосинтеза за вегетацию, г/м2/сут.
По мнению Н.Д. Кумсковой (2004), влияние удобрений на площадь листьев, продуктивность фотосинтеза, прирост сухого вещества гречихи значительное (рис. 61).
Прирост зелёной и сухой массы гречихи находился в тесной зависимости от удобрений. На вариантах без их применения и при внесении азота прирост зелёной массы продолжался до уборки, что важно при выращивании гречихи на зелёное удобрение. Внесение фосфора, совместно с комплексным удобрением, обеспечивало накопление биомассы на 1–2-е декады раньше (табл. 57).

Рис. 61. Влияние удобрений на рост площади листьев, продуктивность фотосинтеза, прирост сухого вещества гречихи по декадам (по Н.Д. Кумсковой, 2004)
Динамика нарастания зелёной и сухой массы гречихи в зависимости от удобрений (по Н.Д. Кумсковой, 2004), т/га
Удобрения, в зависимости от влажности почвы, по-разному влияли на формирование листовой поверхности гречихи (Н.Д. Кумскова, 2004). При выпадении большого количества осадков в начальный период вегетации на варианте с азотом формировалась низкая площадь листьев – 21,7 тыс. м2 на 1 га. На делянках, удобренных только фосфором и при совместном внесении азота, фосфора и калия листовая поверхность была больше на 19,3 и 25,1 тыс. м2 на 1 га, по сравнению с вариантом N40, и на 8,4 и 14,2 тыс. м2 – по отношению к контролю.
Существенно изменялась листовая поверхность в зависимости от густоты посева (Н.Д. Кумскова, 2004). С увеличением нормы высева с 2 до 5 млн. всх. зёрен на 1 га площадь листьев на одном растении снизилась с 175 до 104 см2, а на 1 га увеличилась с 26 до 57 тыс. м2.
Т.М. Русакова (1974) также считает, что площадь листьев гречихи на высоком агрофоне может превышать показатели контроля в 2,0–2,5 раза.
С.У. Броваренко (1970), на основании многолетних исследований в Западной Сибири, пришёл к выводу о том, что лучшее развитие ассимиляционной поверхности листьев гречихи происходит на узкорядных посевах, в сравнении с широкорядными. Площадь листьев на узкорядных посевах достигает 44,4 тыс. м2 на 1 га, на рядовых – 29,8, а на широкорядных – только 19,6 тыс. м2 на 1 га, соответственно, и урожайность на последних ниже. Можно высказать предположение, что урожайность на широкорядных посевах снижалась из-за угнетения культурных растений сорняками, так как известно, что рядовые, и особенно узкорядные посевы, успешно противодействуют засорённости.
С.И. Рак (1967) также считает, что площадь листьев гречихи при рядовом способе посева примерно в 1,5 раза больше в сравнении с листовой поверхностью на междурядьях 0,45 м.
По мнению К.А. Савицкого (1970), большое производственное значение имеют широкорядные посевы гречихи. На чистых от сорняков почвах они эффективнее сплошных рядовых. Листовая поверхность на широкорядном посеве в 1,3–1,4 раза больше, чем на сплошном. Это способствует повышению продуктивности фотосинтеза у растений широкорядного посева в фазе плодообразования в 1,5–1,9 раза.
Наши наблюдения подтверждают данные результаты. В зависимости от ширины междурядий площадь листьев возрастает в 1,4 раза в пользу разреженного посева, увеличение нормы высева также способствует росту ассимиляционной поверхности примерно в 1,5 раза.
Преимущество узкорядного способа посева перед обычным рядовым и широкорядным можно объяснить более слабым ростом, развитием и ветвлением растений в разреженных посевах, что наблюдается при наличии сорняков в междурядьях.
Гречиха интенсивно формирует листья в фазу всходов и бутонизации, к началу цветения их доля составляет 30–50 % от всей биомассы (Г.В. Копелькиевский, 1963). Способы посева и нормы высева значительно влияют на формирование площади листьев. Так, к периоду начала плодообразования размер листовой поверхности быстро увеличивается, но затем скорость данного процесса снижается.
Наши наблюдения показали, что в условиях лесостепи Алтайского края площадь листьев гречихи в фазу плодообразования составляет 48,9–69,8 тыс. м2/га (табл. 58).
Площадь листьев и продуктивность фотосинтеза посевов гречихи
(средняя за 2010–2011 гг.)
Норма высева, млн. всх. зёрен на 1 га
площадь листьев, тыс. м2/га
площадь листьев, тыс. м2/га
площадь листьев, тыс. м2/га
Широкоряд-ный (0,45 м)
Широкоряд-ный (0,60 м)
В связи с тем, что по урожайности зерна гречихи широкорядные посевы 0,60 м уступают таковым с междурядьями 0,45 м, можно предположить, что солнечная радиация и питательные вещества в разреженных посевах используются на создание вегетативной массы, а не зерна.
Таким образом, создавать оптимальную площадь листьев гречихи можно различными технологическими приёмами. Подбор лучших агротехнических условий, обеспечивающих оптимальное развитие листовой поверхности, имеет важное практическое значение.
Главным показателем фотосинтетической деятельности растений является чистая продуктивность фотосинтеза. Связь её с урожаем самая непосредственная и при равенстве площади листьев урожай сухой биомассы растений прямо пропорционален чистой продуктивности фотосинтеза (Т.М. Русакова, 1974; Schilling Herbert, 1980; J.H. Golbeck, 1992).
Продуктивность фотосинтеза гречихи не остается постоянной в течение всего периода вегетации. Наибольшую величину она имеет в период бутонизации-цветения, то есть в период наибольшего прироста сухой массы растений (Т.М. Русакова, 1974). Чистая продуктивность фотосинтеза следует за уровнем минерального питания: наиболее высокие показатели её наблюдаются у растений на чернозёмной почве с внесением минеральных удобрений (7,44 г/м2 в сут. в среднем за вегетацию).
Внесение минеральных удобрений в опытах Т.М. Русаковой (1974) довольно результативно отразилось на величине чистой продуктивности фотосинтеза. Это можно объяснить тем, что хорошее минеральное и, прежде всего, азотное питание активизирует фотосинтез деятельных, хорошо освещённых листьев верхних и средних ярусов, которые в сильной степени поддерживают активную жизнедеятельность листьев нижних ярусов. Последние, в условиях ослабленного света и затруднённого минерального питания, обычно быстро стареют и отмирают. Очевидно, поэтому на высоких фонах минерального питания образуются растения с большой площадью листьев и с высокими показателями чистой продуктивности фотосинтеза.
Анализ результатов определения чистой продуктивности фотосинтеза гречихи в условиях Новосибирской области говорит о том, что изменение этого показателя во времени и по способам возделывания культуры происходит по аналогии с изменением площади листьев и фотосинтетического потенциала. При этом, независимо от способа возделывания, чистая продуктивность фотосинтеза всегда была выше в благоприятные по увлажнению годы (А.Д. Гончаров, 2008).
По мнению Н.М. Городнего (1980), гречиха имеет лучшую, чем многие культуры способность к синтезу органических кислот (в мг на 1 г сухого вещества растения): гречиха – 7,01; нут – 5,08; люпин синий – 4,60; горчица – 4,55; горох – 3,02; овес – 2,88; кукуруза – 1,38. Всё это позволяет считать гречиху нетребовательной к почвам и приводит к тому, что её часто высевают на малоплодородной почве, где другие культуры почти не дают урожая.
По подсчетам А.А. Ничипоровича (1959, 1966), посевы культурных растений в процессе фотосинтеза способны образовывать до 8–10 г сухой биомассы на 1 м2 листовой поверхности, а потенциальная продуктивность может достигать 20–40 г/м2
в сутки. Этот показатель зависит от освещённости солнцем, притока питательных веществ и воды.
Гречиха имеет много точек роста, которые потребляют большое количество продуктов ассимиляции. Ассимиляция проходит при фотосинтезе, который напрямую зависит от величины листовой поверхности и интенсивности солнечного освещения. Величина листовой поверхности гречихи по отношению к количеству цветков на растении составляет всего 0,30–0,56 см2 на один цветок. Эта площадь слишком мала для полного обеспечения цветка пластическими веществами. Если период быстрого роста побегов совпадает со временем развития цветков и плодов, то при недостатке продуктов ассимиляции растущие верхушки стеблей и ветвей оказываются обеспеченными пищей лучше, чем большая часть репродуктивных органов (Н.М. Городний, 1980; J. Oppereer, 1985; S. Wojcik, 1991).
Листья гречихи не обладают высокой энергией усвоения углекислоты. При высокой температуре днём, даже при достаточной влагообеспеченности растений, листья подвядают. Фотосинтез ухудшается из-за быстрого роста вегетативных органов. В результате листья нижнего яруса сильно затеняются и освещаются рассеянным светом, существенно потерявшим физиологически активные лучи. Это снижает образование пластических питательных веществ, недостаток которых вызывает отмирание цветков и завязей (Vong Hguyen Quoc, 1977; И.Н. Елагин, 1984).
Гречиха – светолюбивое растение. Чем лучше освещены её листья, тем энергичнее и больше она усваивает из воздуха углекислоту, перерабатывает её в легкоусвояемые питательные вещества.
В условиях почвенной засухи рост гречихи прекращается, но развитие её продолжается, причем происходит оно быстрее, чем в нормальных условиях. В результате образуются карликовые растения, которые быстро отцветают и созревают, а урожай зерна резко снижается (Н.М. Городний, 1980).
По мнению М.Н. Городнего (1980), на загущенных посевах верхний ярус листьев гречихи поглощает только 60–70 % солнечной радиации. Средний и нижний ярусы получают третью часть и меньше общей энергии солнечного света. В среднем ярусе широкорядного посева проникающая радиация составляет 60–80 % общей падающей, а в нижнем – 50–70 %. Разреженный посев лучше использует световой режим для процессов фотосинтеза, по сравнению с загущенным.
Известно, что при затенении нижних ярусов листьев растений чистая продуктивность фотосинтеза начинает уменьшаться. Лучший по количеству и качеству урожай зерна гречихи можно получить только в посевах, обладающих достаточной по размерам площадью листьев – 40–45 тыс. м2/га при оптимальном ходе её наращивания (М.Н. Городний, 1980). В наших исследованиях лучшей ассимилирующей поверхностью гречихи был показатель на уровне 56,7 тыс. м2/га. Расчёты говорят о том, что в процессе фотосинтеза гречиха может формировать за сутки до 7,34 г сухой биомассы на 1 м2 листовой поверхности. При этом лучшие показатели урожайности отмечались при среднесуточной продуктивности фотосинтеза – 5,83 г сухой биомассы на 1 м2.
Таким образом, сравнение показателей ассимиляционной поверхности и фотосинтетической деятельности растений гречихи между вариантами, позволяет сделать вывод о том, что способы посева и нормы высева оказывают определяющее влияние на урожай зерна. Эти агротехнические приёмы можно отнести к наиболее результативным.
Фотосинтетический потенциал посевов гречихи является обобщающим показателем воздействия многих агротехнических факторов, в том числе способа посева и нормы высева. Н.Д. Кумскова (2004) считает, что фотосинтетический потенциал гречихи может составлять от 1,076 до 1,439 млн. м2 дней /га в зависимости от вносимых удобрений.
По мнению Н.М. Городнего (1980), основная причина отмирания большей части репродуктивных органов гречихи заключается в слабом снабжении их пластическими веществами. Невысокая озернённость гречихи обусловлена неудовлетворительным питанием её цветков из-за низкой облиственности растения и одновременного развития вегетативных и генеративных частей. Продукты синтеза перемещаются в растениях к тем органам, где они потребляются, и в том количестве, в котором они используются. Чем активнее жизнедеятельность организма и чем он мощнее, тем большее количество продуктов ассимиляции поступает в растение. Если образуется много растущих органов и общая потребность их в органическом веществе превысит синтетическую деятельность ассимиляционного аппарата растения, у такого растения наблюдается недостаток продуктов ассимиляции и его растущие части голодают. При этом больше страдают органы, которые растут менее интенсивно, в которые органические вещества поступают слабее. Их рост приостанавливается, но если недостаток питания длителен или даже постоянен, растения гибнут.
Бороздковые посевы гречихи в условиях Новосибирской области, в сравнении со сплошными рядовыми, формировали более значительный фотосинтетический потенциал. При этом преимущество бороздковых посевов по данному показателю последовательно нарастало от ранних периодов роста и развития растений, к более поздним. В среднем за 3 года исследований А.Д. Гончарова (2008), за период всходы-бутонизация, фотосинтетический потенциал бороздковых посевов был больше соответствующего показателя сплошных рядовых на 15 %, а за период бутонизация-цветение – на 28 %, за период цветение-созревание – на 30 %.
В опытах Н.Д. Кумсковой (2004) высокая облиственность растений и урожайность гречихи отмечены на посевах нормой 3 млн. всх. зёрен на 1 га. При высеве 2 млн. семян снижался выход зерна из-за недостаточной густоты стояния растений, что приводило к неэффективному использованию пашни. Нормы высева 4 и 5 млн. зёрен нерациональны, так как в таких посевах формировалась избыточная вегетативная масса. Наиболее высокая масса листьев гречихи, по отношению к массе растений (облиственность), наблюдалась в фазу бутонизации. Затем облиственность растений гречихи к началу цветения снижалась в два раза, достигала минимального значения в фазу созревания. На делянках, удобренных азотом, снижение её было более интенсивным, чем на контроле и других фонах. В этом варианте облиственность к моменту созревания равнялась 13,3 %, вместо 18,6 % на контроле.
Полученные нами данные для лесостепной зоны Алтайского края не противоречат результатам исследований Н.Д. Кумсковой (2004).
Наши наблюдения говорят о том, что максимального значения фотосинтетический потенциал достигал в разреженном травостое в фазу плодообразования, то есть при широкорядном способе посева (табл. 59).
Фотосинтетический потенциал и облиственность гречихи посевной
(средние за 2010–2011 гг.)
Читайте также: