
Что появилось у покрытосеменных растений в процессе эволюции побеги с листьями проводящие клетки
Обновлено: 07.07.2024
Можно проследить следующие главные направления эволюции этой структуры:
1. От большого и нефиксированного числа частей к малому и четко определенному.
2. От четырех типов частей до трех, двух, а иногда и до одного у более продвинутых эволюционно групп. Побег укорачивается и их исходное спиральное расположение частей перестает замечаться. Части цветка срастаются - приспособление к опылению насекомыми.
3. От верхней завязи к нижней. Лучшая защита семяпочки.
4. От актиноморфности к зигоморфности.
5. Переход от одиночных цветков к соцветиям, размеры, яркость привлекают насекомых.
6. Приспособление к опылению ветром обусловило редукцию околоцветника и образование голых цветков.
7. Появление однодомности и двудомности у ветроопыляемых растений обеспечило перекрестное опыление.
Сочетание всех этих признаков у разных групп неодинаково. Цветки одних могут быть более сложными по одним признакам, а по другим примитивные. Семейство Крестоцветные - 4-х членный цветок имеет один пестик - признак высокой организации, завязь всегда верхняя - признак простой организации.
Судить об эволюционной продвинутости нужно по нескольким признакам.
Эволюционный успех цветковых растений в значительной степени был обусловлен их параллельным развитием с различными животными. Они оказывали друг на друга давление и во многом определили эволюцию своих партнеров. Яркая окраска цветков, душистый аромат, съедобная пыльца и нектар - все это средства для привлечения животных. Адаптация цветка была направлена на максимальное увеличение шансов для переноса пыльцы насекомыми. Это опыление было надежно.
Важным признаком успеха эволюции покрытосеменных является биохимическая эволюция. В некоторых группах выработалась способность образовывать вторичные метаболиты (алколоиды, хитоны, эфирные масла, флавоноиды и др.) - ядовитые для животных вещества, защищающие растения от большинства фитофагов (вредители).
К группам с наиболее специализировавшимися в ходе эволюции цветками принадлежат сложноцветные (Compozitae) среди двудольных (Dicotyledones) и орхидные (Orchidaceae) из однодольных (Monocotyledones).
Рис. 184 Стадии эволюции тычинок (микроспорофиллов) (1—6) от примитивного микроспорофилла дегенерии фиджийской (Dеgеneria vitiensis) (1)до высоко-специализированного типа (6)
Рис. 185 Стадии эволюции плодолистика (1-6)от примитивного кондупликатного плодолистика типа дегенерии фиджийской (Dеgеneria vitiensis) (1) до специализированного типа с резко дифференцированным столбиком и головчатым рыльцем (6)
Рис. 186 Схема эволюции основных типов гинецея. Паракарпный и лизикарпный типы изображены в двух стадиях эволюции, что дает представление о двух разных путях их происхождения от исходного от апокарпного типа. По многих случаях паракарпный гинецей происходит непосредственно от апокарпного.
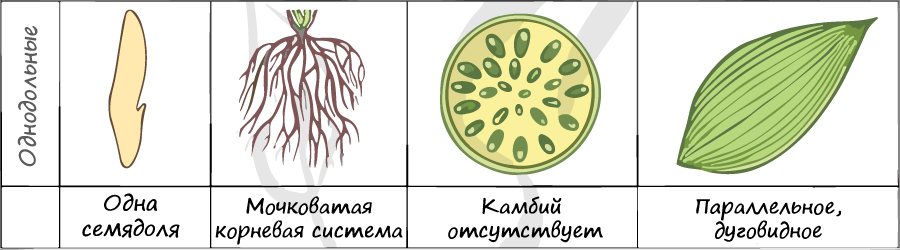
3. Отличительные признаки классов двудольных и однодольных
Таблица 18. Отличительные признаки классов цветковых растений
| Признак | Однодольные | Двудольные |
| Строение семени | Одна семядоля. | Две, редко 3,4 семядоли. |
| Строение цветка. | Число частей 3 или кратное 3. | Число частей по 4-5 или кратное 4. |
| Форма листа. | Простые, цельнокрайние, обычно сидячие. | Простые или сложные, края рассеченные. |
| Жилкование листа. | Параллельное или дуговое. | Сетчатое, перистое, пальчатое. |
| Стебель. | Травянистый. | Травянистые или деревянистые. |
| Проводящие пучки. | Проводящие пучки разбросаны по стеблю и не имеют камбия. Закрытые. | Проводящие пучки в центре стебля расположены по кругу, имеют камбий. Открытые. |
| Корневая Система. | Мочковатая. | Стержневая. У некоторых мочковатая. |
По данным палеонтологии предки покрытосеменных появились в начале мелового периода и очень быстро заселили большие пространства суши. В этот период на земном шаре произошло резкое изменение климата: уменьшилась влажность, увеличилось освещенность и сухость воздуха. Большая часть голосеменных, господствовавших в то время, не смогла приспособиться к новым условиям (к сухости) и вымерла за исключением хвойных, которые сохранились благодаря небольшой площади листовой поверхности. Покрытосеменные оказались лучше приспособленными к новым условиям, более пластичными. Они начали бурно развиваться, дали огромное разнообразие форм и заняли господствующее положение.
Самые древние покрытосеменные были, вероятно, древесными растениями с небольшими моноподиально ветвящимися стволами и немногочисленными толстыми ветвями. От них произошли более крупные формы с сильно разветвленной кроной и симподиальным ветвлением, позже появились кустарники, кустарнички, многолетние травы, и наконец, однолетние травы.
4. Причина процветания покрытосеменных растений
При сравнении жизненных циклов высших растений наиболее эволюционно продвинут у покрытосеменных. Сложно устроен спорофит (бесполое поколение). Гаметофит представляет несколько клеток. Женское половое поколение полностью развивается на бесполом поколении под защитой пестика. Опыление ветром и насекомыми еще более надежно обеспечивает перенос пыльцы, а следовательно, и оплодотворение яйцеклетки. Разнообразие в строении вегетативных органов, образовании семян и плодов способствовали расселению покрытосеменных в самые различные условия обитания.
5. Филогенетическая система покрытосеменных
Все семейства и порядки покрытосеменных растений находятся между собой в более или менее близком родстве. Филогенетические системы растительного мира обычно изображают схематично в виде родословного дерева, на котором отдельные направления эволюции растений показаны как его боковые ветви. Система покрытосеменных А.А.Гроссгейма изображена на схеме (прилагается) в виде родословного дерева, но в плане, как проекция сверху. На этой схеме самые древние порядки, являющиеся родоначальниками остальных покрытосеменных, находятся в центре. Таковы порядок Лютикоцветные, представленный главным образом травянистыми растениями, и порядок Магнолиецветные, представленный только древесными растениями.
На схеме порядки изображены кружками, величина которых приближенно отражает число видов в порядке. Порядки, в которые входят древесные породы, отмечены черным кружком. Академик А.А.Гроссгейм различает три ступени в развитии цветка покрытосеменных и границы этих ступеней отмечает на схеме двумя окружностями. К первой ступени развития отнесены порядки с наиболее просто устроенными цветками: с неопределенным числом частей цветка, со спиральным их расположением, с многочисленными пестиками, с верхней завязью. Эти порядки помещены внутри меньшей окружности, что подчеркивает их близость к древним формам.
Ко второй ступени развития отнесены порядки с более сложным строением цветка. Число частей в цветке уменьшается до 3-5, расположены они чаще кругами. Эти порядки в схеме расположены между внутренней и внешней окружностью.
К третьей ступени развития отнесены порядки с ярко выраженной приспособленностью цветков к опылению определенного типа, что усложнило их строение. Растения группы насекомоопыляемых имеют сросшийся неправильный венчик, нижнюю завязь. Ветроопыляемые растения отличаются вторичным упрощением цветка - редукцией венчика, верхней завязью. Эти порядки в схеме расположены за пределами второй окружности.
От порядка Лютикоцветные берет начало несколько направлений эволюции. Влево ведет линия развития к порядку розоцветные. В порядке наблюдается усложнение строения цветка в виде перехода к одному пестику и нижней завязи. Порядок Розоцветные очень разнообразен, и разные его представители дают начало нескольким направлениям эволюции. Одно из них ведет к крупному порядку Бобовоцветные, другое - к порядку Зонтикоцветные. У них усложнение цветка выразилось в переходе к нижней завязи, уменьшении числа частей цветка и в сильной рассеченности пластинки простого листа.
Развитие древесных форм от порядка розоцветные привело к образованию порядка Букоцветные с переходом у них к простому листу, ветроопыляемости, полной редукции околоцветника.
От розоцветных родственные связи ведут к большому порядку Трубкоцветные, у которых наблюдается переход к супротивному листорасположению, неправильному околоцветнику, срастанию венчика, уменьшению числа тычинок до двух, но завязь всегда остается верхней.
От Лютикоцветных берет начало отдельная линия эволюции, которая привела к порядку Макоцветные. У них произошло уменьшение числа тычинок от неопределенного и большого к определенному и небольшому, уменьшение числа частей околоцветника и образование одного пестика.
По этому же пути пошло развитие более высокоорганизованного порядка Верескоцветные, которые характеризуются простыми листьями, переходом к сросшемуся венчику, к нижней завязи и 4-5-членному цветку.
Заканчивает эту линию развития большой порядок сложноцветные. Этот порядок отличается типом соцветия - корзинкой. В цветке наблюдается редукция чашечки, срастание венчика и пыльников пяти тычинок, образование одного пестика, появление нижней завязи.
Линия развития от Лютикоцветных привела к порядкам Центросеменные и Гречихоцветные, которые характеризуются переходом к супротивным листьям и срастанию частей околоцветника, числом тычинок, равным числу частей околоцветника или удвоенным, 3-5-членным цветком. Наряду с насекомоопыляемыми появляются и ветроопыляемые виды.
В порядки Лилиецветные, Осокоцветные, Злакоцветные относятся травы, отличающиеся строением вегетативных органов. Листья линейные или ланцетные, с паралельным или дуговидным жилкованием, цветки трехчленные, с двойным простым или редуцированным околоцветником, тычинок 3-6, пестик 1. В этом ряду наблюдается переход от насекомоопыления к ветроопылению.
Система А.А.Гроссгейма дает наглядное представление о родственных связях и путях эволюции различных порядков цветковых растений.
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру: от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.

Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) - важнейшими производителями органических веществ на суше, как водоросли - в морях и океанах.
Цветок - генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения. Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище - завязи, сформированной из плодолистика (-ов).
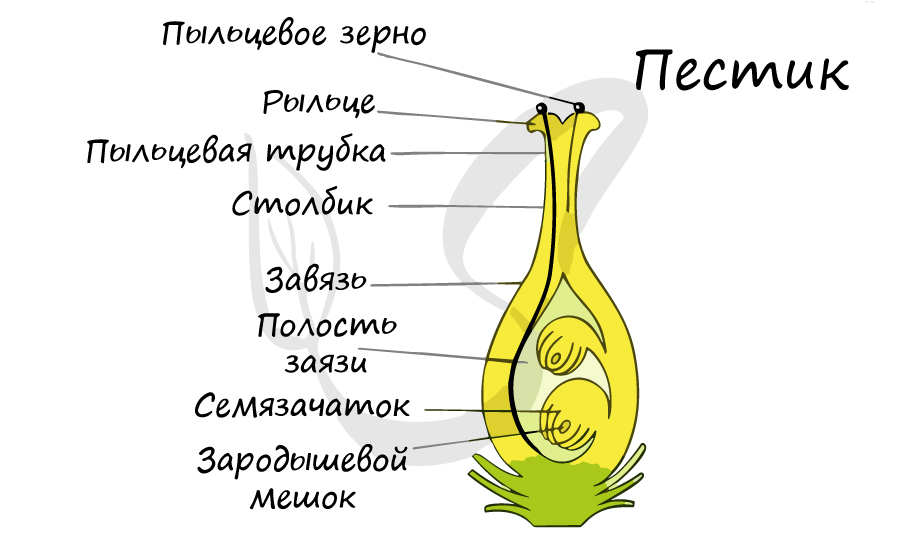
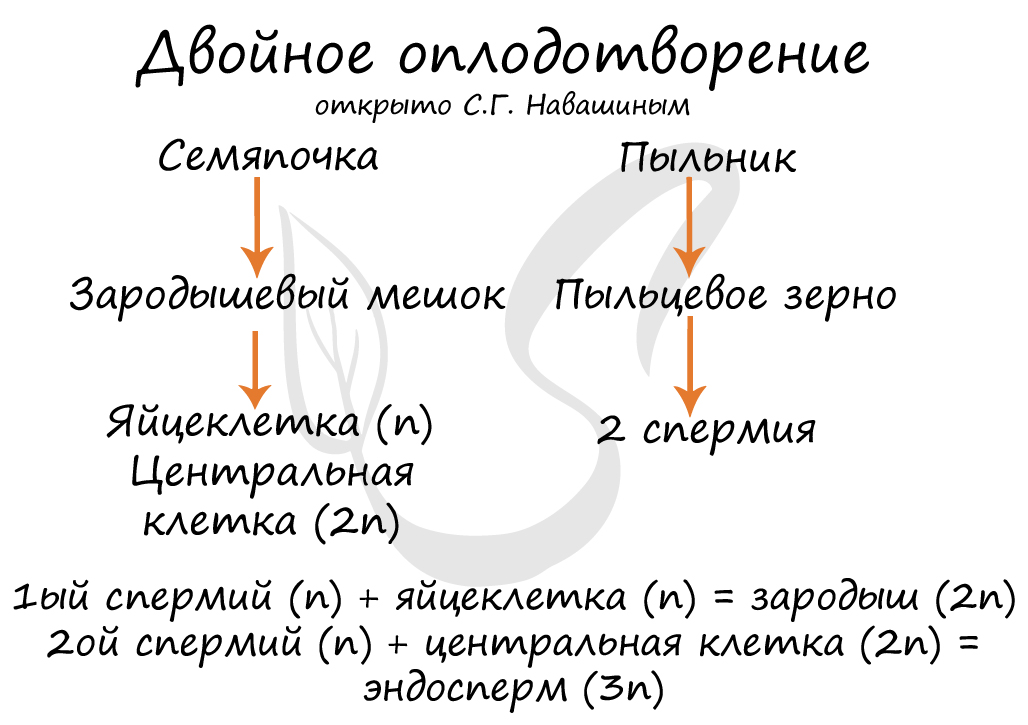
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества - эндосперма (3n). Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод - генеративный орган, служащий для защиты и распространения семян.
Ксилема - проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами, а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

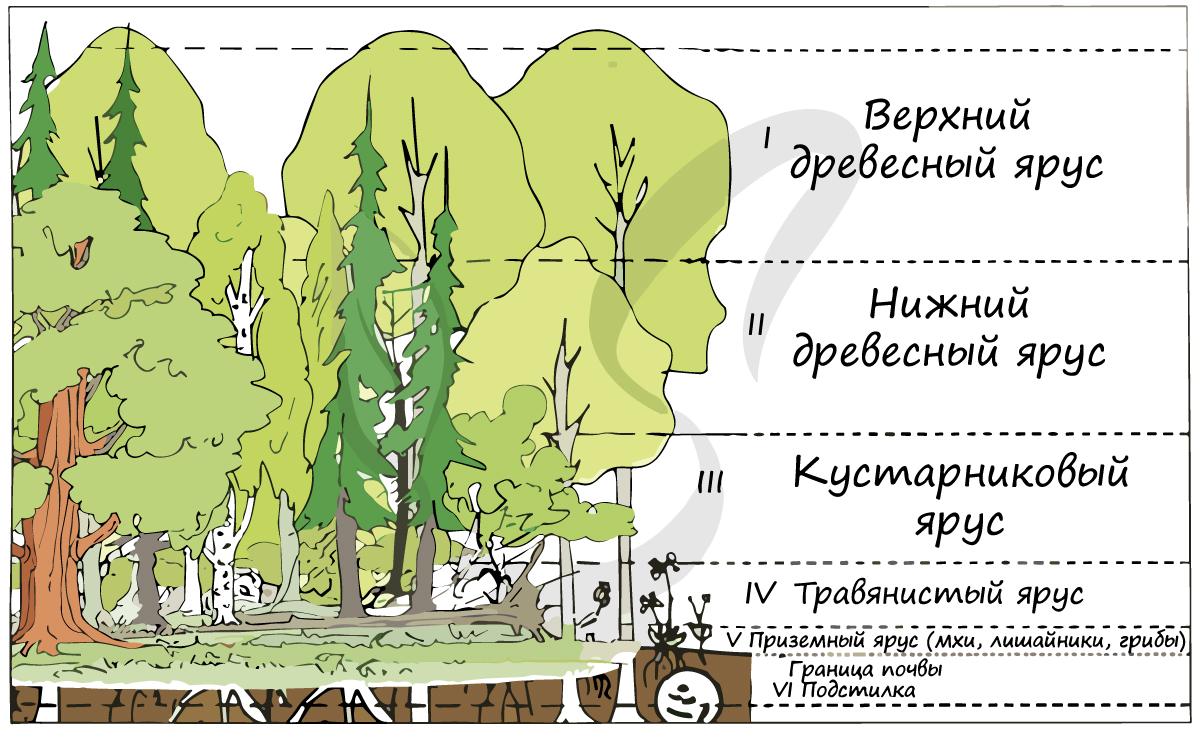
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых :)
Классы покрытосеменных
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные, бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
-
Двудольные - семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
-
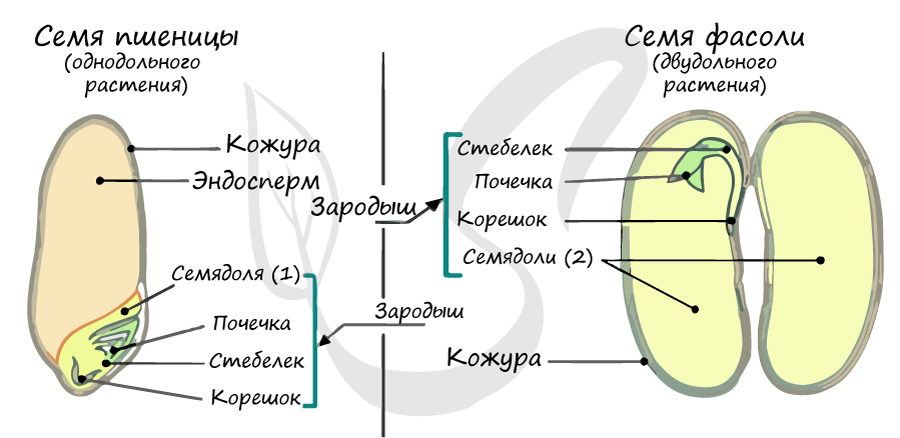
В составе зародыша обычно имеется две семядоли
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни. Главный корень развивается из зародышевого корешка.
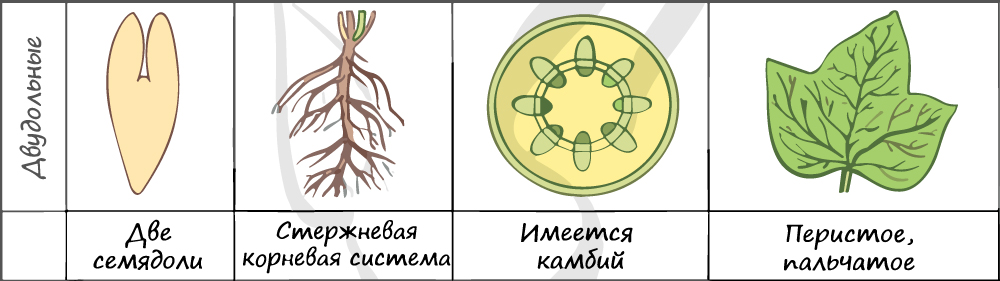
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

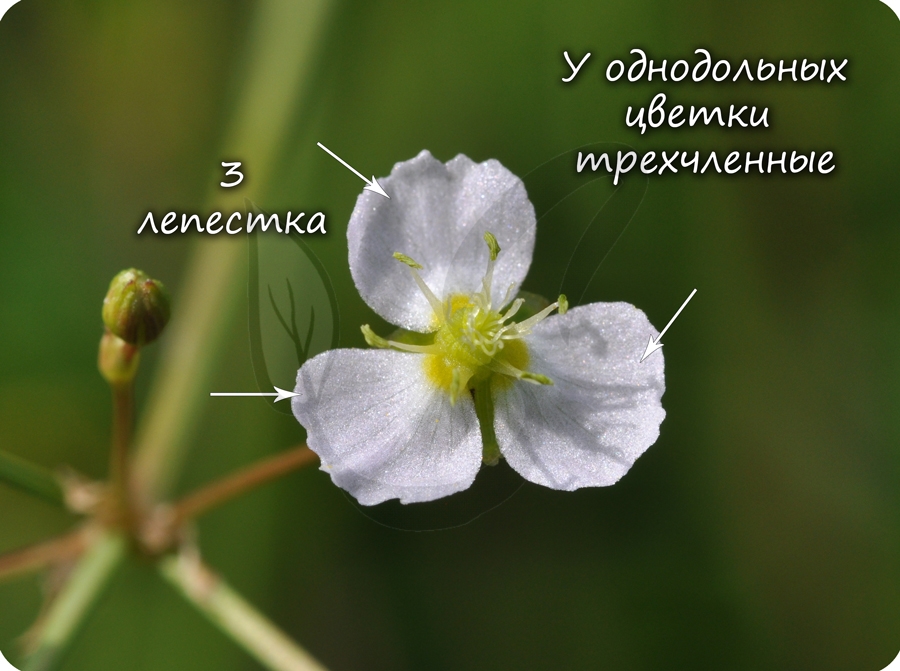
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
Эндосперм семени
Эндосперм (от греч. endon - внутри + греч. sperma - семя) - запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у - березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
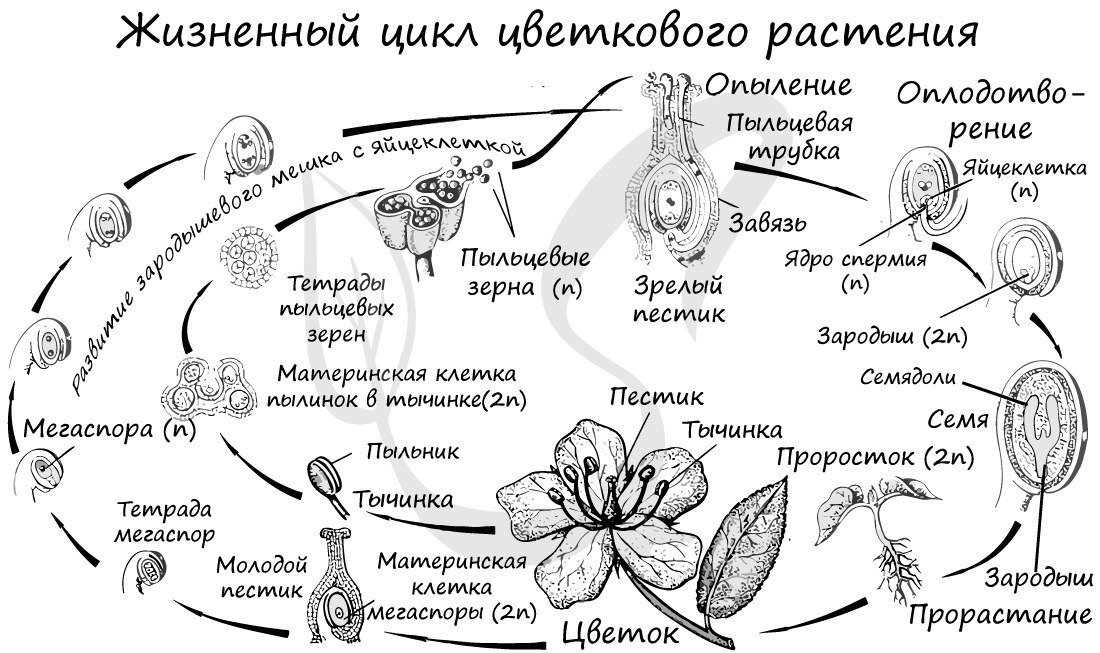
Жизненный цикл
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n) в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) - мужской гаметофит. В завязи пестика в семязачатке формируется женский гаметофит - зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток. Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) - запасного питательного вещества. Другой спермий (n) сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник - образуется плод. Своим внешним видом плоды привлекают животных, и те их охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия, они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте - почти все культурные растения принадлежат к этому отделу! Цветковые имеют медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Свидетельство и скидка на обучение каждому участнику
Основные направления эволюции цветка у покрытосеменных растений.
Появление покрытосеменных, или цветковых, растений представляет собой один из самых значительных этапов эволюции органической жизни на Земле. Покрытосеменные являются господствующей группой растений на нашей планете. Цветковые, указывает Козо-Полянский, имеют поистине неизмеримое значение в природе - значение посредника между веществами и существами, между природой минеральной и органической. Не будь этого посредничества, и животная жизнь не могла бы развернуться в ту грандиозную картину, которую мы видим.
Сходство цветка со стробилами хвощей и плаунов и со спорофиллами папоротников является чисто внешним, т.к. в цветке совмещены оба - половое и бесполое - поколения. Т.е. женский гаметофит гомолог зародышевого мешка, а мужской гаметофит - пыльцевого зерна, содержащего вегетативную и генеративную клетку. То, что принимается за цветок у папоротников, хвощей и плаунов, как известно, представляет собой орган бесполого размножения, развивающийся на спорофите. Намного ближе по своей сути цветок покрытосеменных к стробилам голосеменных, но и здесь есть значительные отличия. Он очень сильно специализирован благодаря далеко зашедшим эволюционным процессам. Истинная природа органов цветка с трудом распознается и нередко вызывает различные толкования
Строение цветка
Цветок имеет ось, или цветоложе , которая несет листочки околоцветника , тычинки и пестики , состоящие из одного или нескольких плодолистиков . В завязях пестиков содержатся семязачатки (семяпочки). После опыления и оплодотворения завязи превращаются в плоды, а семязачатки - в семена. Плод не может возникать независимо от цветка.
Цветок бывает верхушечным или выходит из пазухи кроющего листа, большей частью отличающегося по форме от вегетативных листьев. Участок между кроющим листом и цветком называют цветоножкой . На цветоножке располагаются также два или один маленьких листочка, которые называют предлистьями или прицветничками.
В зависимости от наличия или отсутствия околоцветника, а также от его строения, различают следующие типы цветка :
1. Гомохламидный . Околоцветник простой, т.е. листочки его примерно одинаковые, чаще спирально расположенные, в довольно большом числе, чашечковидные или венчиковидные (магнолия, купальница, лилия); характерен для более примитивных семейств покрытосеменных.
2. Гетерохламидный. Околоцветник двойной, т.е. дифференци-рованный на чашечку и венчик (гвоздика, колокольчик, горох).
3. Гаплохламидный , или монохламидный . Лишь один круг листочков околоцветника, чаще чашечковидные (крапива, вяз, лебеда).
4. Апохламидный. Оклоцветника нет, цветки голые (ясень, ива). В настоящее время полагают, что в подобных случаях имела место утрата околоцветника.
Вопрос о становлении цветка в процессе эволюции занимает центральное место в проблеме происхождения покрытосеменных. Повышенный интерес филогенетиков к цветку находит объяснение. Б.М. Козо-Полянский указывает, что для филогении признаки тем ценнее, чем они больше говорят о прошлой истории и о предках растения: признаки вегетативной сферы несравненно пластичней признаков цветка. В свое время Имс также подчеркивал, что в цветке, именно в особенностях проводящей системы, обнаруживается консервативность: 1) скелет цветка обладает консервативностью, и 2) консервативность генеративных органов. Вопросы о происхождении цветка, о его первичном типе, о природе его частей являются поэтому органической и существенной частью проблемы происхождения покрытосеменных
Прототипы цветка
Красилов считает, что цветкоподобные структуры могли развиться на различной морфологической основе, и прототипы цветка можно отнести к трем категориям:
1. Антокормоиды . Они имели укороченный побег с верхушечным собранием пыльцевых и семенных органов среди неизменных листьев. Примером могут служить брахибласт Гингко, апикальная зона которого, окруженная наружными толстыми и внутренними тонкими брактеями, продуцирует пыльцевые стробилы. Есть основание полагать, что семенные и пыльцевые органы кейтониевых, чекановскиевых и диропалостахиевых были собраны таким же образом.
2. Многоузловые и одноузловые антостробилы . Многоузловый антостробил Гнетовых несет в каждом узле мутовчатые собрания пыльцевых и семенных купулятивных органов, поддерживаемых брактеями. Одноузловой антостробил беннетитовых в типичном случае представляет собой собрание коротких овулифоров, перемежающихся с межсеменными чешуями.
3. Предцветки, или протантоиды . Предцветок Баисии состоит из апикальной купулятной семяпочки на ширококоническом рецептакуле в окружении брактей с длинными пучками волосков
Теории происхождения цветка
Огромное разнообразие цветков покрытосеменных и их резкое отличие от соответствующих органов голосеменных порождают значительные трудности в объяснении происхождения цветка.
Одна из известнейших теорий происхождения цветка - псевдантовая теория Веттштейна - имеет в настоящее время лишь историческое значение.
Веттштейн полагал, что среди всех покрытосеменных простейшими и наиболее примитивными цветками обладают так называемые однопокровные, куда относятся буковые, березовые, вязовые и другие семейства. Здесь сохраняется раздельнополость цветков, характерная для стробилов голосеменных. В качестве примера он приводит эфедру, которая имеет собрания стробилов. Каждое собрание состоит из короткой оси, несущей 2-8 супротивных пар чешуевидных кроющих листьев, в пазухах которых сидит по одному просто устроенному микростробилу.
Мужской цветок австралийских казуарин можно вывести из собраний микростробилов эфедры, если предположить некоторые модификации в покровах цветков.
Вторая ступень характеризуется увеличением числа тычинок. На третьей ступени возникает венчик из андроцея, а существовавший до того околоцветник становится чашечкой.
Весьма существенным положением теории Веттштейна является то, что на третьей ступени мужской цветок превращается в обоеполый. Поскольку посещение насекомых только тогда полезно растению, когда они соприкасаются не только с тычинками, но и с рыльцем. Отбор фиксировал случаи, когда в одном цветке появлялись и тычинки и плодолистики. Обоеполый цветок является, таким образом, сложным синтетическим образованием.
Из всего сказанного вытекает, что наиболее примитивные покрытосеменные, с точки зрения Веттштейна, имеют просто устроенные цветки с простым околоцветником или без него, часто раздельнополые
Теломная теория. Теломная теория возражает против основных понятий классической морфологии о природе частей цветка, как метаморфизированных листьев. Предпосылками для обоснования теломной теории явилось открытие в 1917 - 1920 гг. псилофитовых, как особой группы наиболее примитивных высших растений, по-видимому. Давшей начало папоротникообразным, и, стало быть, голо- и покрытосеменным. К простейшим псилофитам относятся роды Риния, Хорнеофитон и Коксония.
Знакомство с псилофитовыми сделало очевидным, что лист не является исходным органом высшего растения, а выработался в процессе длительной эволюции. Спорангий же филогенетически старше.
Немецкий ботаник Циммерман полагает, что все органы высших растений развились из простых органов - теломов, представляющих конечные веточки дихотомической системы ветвления псилофитовых. В процессе эволюции теломы срастаются в синтеломы, стерильные и фертильные. В дальнейшем стерильные синтеломы дифференцируются на листья и оси побега, а фертильные преобразуются в спорофиллы, которые дают начало цветку.
Некоторые особенности беннеттитовых - обоеполость их стробилов, расположение частей, форма оси - заставили предполагать их родство с многоплодниковыми.
Предполагается, что в процессе эволюции редуцируются микроспорофиллы и число микроспорангиев сокращается до четырех. Открытые мегаспорофиллы, смыкаясь краями и срастаясь, превращаются в замкнутые плодолистики с расположенными внутри семязачатками. Пыльца начинает улавливаться плодолистиками, возникает антостробил, т.е. образование, которое уже является цветком, но в то же время ясно обнаруживает стробилярную природу.
Среди современных покрытосеменных встречаются растения, обладающие теми примитивными чертами, что и гипотетическая модель Арбера и Паркина, а в некоторых случаях и еще более примитивными.
Вряд ли может быть обнаружен как среди ныне живущих, так и среди ископаемых растений цветок, полностью соответствующий арберовскому прототипу, т.к. стадии развития отдельных органов не совпадают во времени. Даже во многих современных цветках можно обнаружить черты как примитивной, так и развитой организации. Это явление носит название гетеробатмия.
Гипотетическому антостробилу наиболее близки цветки магнолиевых, кувшинковых и некоторых однодольных.
В настоящее время положение этой теории было поколеблено, т.к. трудно объяснить некоторые особенности стробилов беннеттитовых. Так, непонятно, почему микроспорофиллы находят исключительно во вдоль сложенном состоянии, почему эти органы располагаются всегда кольцом (а не спирально), почему микроспорофиллы никогда не встречаются вместе с мегаспорофиллами.
Однако нельзя считать, что основы стробилярной гипотезы оказываются окончательно отброшенными. В основе ее лежат представления о цветке как стробиле, обладающем осью - цветоложем и различными органами листового происхождения. Но, с современной точки зрения, плодолистики и тычинки не являются видоизмененными вегетативными листьями, а происходят от соответствующих мега- и микроспорофиллов древних голосеменных - предков покрытосеменных, а те в свою очередь, от спорофиллов папоротникообразных. Черты сходства в строении споролистиков и вегетатативных листьев объясняются параллелизмом
Важно понять, каким образом появился гермафродитизм, т.е. цветок стал обоеполым. В современной группе голосеменных Gnetales, некоторые виды имеют стерильные семязачатки в функциональных мужских репродуктивных частях (их называют эктопические семязачатки). Они выделяют нектар, чтобы привлечь насекомых к мужским органам, в то время как фертильные семязачатки привлекают насекомых к женским структурам, допуская опыление насекомыми. Если стерильные семязачатки можно обнаружить у далеко родственных Gnetales, значит, возможно, они имелись и в мужских структурах проангиосперм.
В соответствии с этой теорией можно предположить, какими особенностями должен был обладать предок покрытосеменных:
1.Иметь мужскую структуру - микроспорофилл, плоский и широкий, без глубоких долей и не сложный, такой, чтобы при наличии семязачатка он становился плодолистиком.
2.Микроспорофиллы должны рождаться вместе, один за другим вдоль специализированного побега, таким образом, чтобы верхние микроспорофиллы могли развиться в плодолистики, а базальные остались мужскими органами и стали тычинками обоеполого цветка.
3.Микроспорофиллы должны были иметь многочисленные жилы, давшие начало нескольким проводящим пучкам в плодолистиках покрытосеменных.
4.Кроме того, предок должен был иметь семязачаток, расположенный эктопически среди микроспорофиллов.
Другая же группа ископаемых Птеридосперм, Corystospermales имеет вещественное доказательство мужской структуры и соответствует данной теории. У представителей этой группы Pteruchus и Pteroma мужские органы от рождения плоские и бездольчатые. У Pteruchus ясно видно, что индивидуальные микроспорофиллы находятся на стволе. Также он имеет многочисленные жилы на стволе, а Pteroma заметного жилкования не имеет. Кроме того группа Corystospermales имела и другие свойства, сходные с тычинками цветковых растений. Например, у большинства цветковых микроспорангии сливаются вдоль по парам, а после созревания открываются, образуя длинную щель. У Pteruchus тоже каждый микроспорангий открывается длинной щелью. И если они будут сливаться в пары, то станут похожи на слившиеся микроспорангии покрытосеменных.
Другой мало известный ископаемый Кретацеоус имел парные микроспорангии с открытой щелью, или единственную пару микроспорангиев на микроспорофилле. Но неясно, относилось ли это растение к Corystospermales.

Эволюция растений
Эволюция царства растений началась с архейской эры (около 3500 млн. лет назад). В этот период на Земле отмечено появление синезеленых водорослей. Эти водоросли принадлежат к группе цианобактерий, поскольку в их клетках отсутствует оформленные ядра|ядра. Таким образом, их можно отнести к прокариотам (доядерным организмам). Среди синезеленых водорослей были одно- и многоклеточные организмы, имеющие возможность осуществлять фотосинтез. Благодаря процессу фотосинтеза, в атмосферу нашей планеты начал|начал поступать кислород, необходимый для жизнедеятельности аэробов.
Позднее|Позднее в протерозойской эре (около 2600 млн. лет назад) Землёй завладели красные и зелёные водоросли. Их господство распространилось и на палеозойскую эру (примерно 570 млн. лет назад). Только в поздний палеозой (силурийский период) отмечена жизнедеятельность на планете древнейших высших растений – риниофитов, или псилофитов. У этих растений были побеги|побеги, но отсутствовали корни и листья. Размножение риниофитов происходило спорами. Они обитали на суше или частично в воде.В существовании нашей планеты новая эра началась с появления высших, или наземных растений. Около 400- 360 млн. лет назад в девонском периоде палеозойской эры на фоне преобладания на Земле риниофитов и водорослей появились первые папоротникообразные (папоротники, хвощи, плауны) и моховидные растения. Они относятся к высшим споровым растениям. Благодаря распространению растений на суше, появились и новые наземные виды животных. Сочетанное изменение в ходе эволюции форм растений и животных обусловило огромнейшее разнообразие жизни на Земле. Облик планеты изменился коренным|корённым образом. Прикреплённый образ жизни растения на суше привёл к появлению расчленения растения на корень, стебель и лист, а также к возникновению опорных тканей и сосудистой проводящей системы. Самые первые наземные растения были маленьких размеров. Они поглощали воду посредством ризоидов, как сохранившиеся по сей день на Земле мхи. В цикле их развития преобладало гаплоидное поколение (гаметофит). Постепенно появлялись более крупные формы растений — папоротникообразные, у которых образовались сложные специализированные органы|органы – корни с корневыми волосками. В цикле развития этих растений на первый план выступает диплоидная фаза — спорофит, являющийся непосредственно самим растением, тогда как гаметофит – это заросток, который выглядит как клубенёк у хвощей и плаунов и как небольшая пластинка в форме сердца|сердца у папоротников. Так осуществлялся постепенный переход от гаплоидного поколения к более совершенному|совершённому — диплоидному. В палеозойскую эру папоротникообразные были громадными растениями, господствовавшими на суше. Однако для их размножения была необходима вода, что ограничивало территорию их существования местностями с повышенной влажностью.
В каменноугольном периоде, который длился с 360 до 280 млн. лет назад, доказано появление на нашей планете семенных папоротников, которые в дальнейшем стали родоначальниками всех голосеменных растений. В это время полностью исчезают риниофиты из-за невозможности конкурировать с более развитыми|развитыми|развитыми растениями. А господствовашие тогда огромнейшие древовидные папоротникообразные после отмирания образовали залежи каменного угля.
В следующем пермском периоде палеозойской эры на Земле появились древние голосеменные растения. Древовидные папоротникообразные постепенно вымирают, а им на смену приходят семенные и травянистые папоротники, завладевая сушей. Особенностью голосеменных растений является то, что размножение их осуществляется семенами|семёнами, не имеющими защиты в виде стенок плода, так как цветков и плодов эти растения не образуют. Половое размножение этих растений осуществлялось независимо от капельно|капельно-водной среды|среды. А их появление в ходе эволюционных метаморфоз было обусловлено перепадами влажности и температуры и изменением рельефа Земли|Земли вследствие поднятия суши|суши, то есть появления горных массивов.
Мезозойская эра наступила около 240 млн. лет назад. В триасовом периоде мезозоя появились современные голосеменные, а в юрском периоде зародились первые покрытосеменные растения. Но господствующие позиции на планете сохранились за голосеменными растениями. Это эра вымирания древних папоротникообразных, не выдерживающих естественного отбора. В процессе появления покрытосеменных растений произошла череда ароморфозов. Во-первых, сформировался цветок – трансформированный побег, приспособленный для образования спор и гамет. Опыление, оплодотворение и образование зародыша и плода происходило непосредственно в цветке. Во-вторых, для лучшей защиты и распространения семена|семёна покрытосеменных растений были окружены околоплодником. Для этих растений характерно половое размножение. К покрытосеменным относятся травянистые растения, деревья и кустарники. Разнообразные видоизменения вегетативных органов|органов (корня, стебля, листа) отмечаются у разных видов растений. Эволюционные изменения покрытосеменных растений происходили за относительно короткий срок, поэтому для них свойственна высокая эволюционная пластичность. Огромное значение в протекании эволюционных преобразований сыграли насекомые-опылители. Покрытосеменные растения более продуктивно осваивают окружающую среду|среду и завоёвывают новые территории, благодаря своим особенностям строения и способности формировать сложные многоярусные сообщества.
В кайнозойской эре, наступившей приблизительно 70 млн. лет назад, на нашей планете стали господствовать существующие и в настоящее время покрытосеменные и голосеменные растения, тогда как высшие споровые растения регрессировали.
Сейчас на Земле произрастают более 350 видов растений, среди них встречаются цветковые, мохообразные растения, папоротники, водоросли.
Эволюция растений

Здравствуйте друзья! Сегодня хотелось бы поговорить о доисторических растениях, о том как они эволюционировали в современные растения.
В растительном мире сегодня преобладают цветковые растения, но плауны и папоротники покрывали Землю в доисторическое время.
Эволюция растений.
Более 400 000 видов флоры известны сегодня, и все они произошли от нескольких древних морских растений. Виды, исчезнувшие с лица|лица Земли|Земли, не входят в это число, поскольку к меняющимся условиям на Земле адаптироваться они не смогли, или не выдержали конкуренцию со стороны вновь появившихся растений, которые лучше были приспособлены к новой среде обитания.
Палеоботаники установили распределение растительного покрова|покрова по поверхности Земли|Земли в разные геологические периоды, а также закономерности его смены. В том, что у растений нет твёрдого скелета, который легко превращается в окаменелость, заключается сложность исследований.
К счастью, иногда можно обнаружить ранние формы флоры в древних илистых отложениях, а в горных породах были найдены некоторые останки растений, их возраст составляет около 3,1 млрд. лет.
О том, что жизнь на планете должна была начаться с появления растениеподобных организмов, ставших важным звеном в пищевой цепи животных в дальнейшем, свидетельствуют окаменелости.
Но намного значимее роль растений в эволюционной истории Земли|Земли, так как они фактически преобразовали атмосферу нашей планеты и сделали её пригодной для существования животного мира.
Вероятно, в условиях изначального содержания огромного количества углекислоты|углекислоты в атмосфере, животные не смогли бы дышать. Углекислый газ в кислород растения преобразуют в процессе фотосинтеза, насыщая им атмосферу.
Основой пищевой цепи послужила способность растений использовать солнечный свет для производства сложных органических веществ. Эволюцию плотоядных и травоядных животных обеспечили растения.
Эволюция, однако, крайне медленный процесс, и естественный отбор благоприятствует особям, приспосабливающимся к изменениям среды|среды обитания, а не просто к изменениям как таковым.
Без воды|воды не могли обходиться древнейшие виды растительного мира, так как структуры необходимые для жизни на суше, у них отсутствовали.
Первые вышедшие растения из воды, вероятно, обосновались в болотах, где их нижняя часть могла постоянно находиться под водой. Скорее всего, первые истинно наземные растения оставались влаголюбивыми и росли возле воды|воды.
Влажная среда для размножения по-прежнему, была необходима печёночникам, мхам и папоротникам, которые развивались как растения с давних времён.
Геологическую колонку с эрами, периодами, эпохами и их длительностью можно посмотреть здесь.
Предшественники цветковых растений – голосеменные, среди них хвойные деревья – нуждались в ветре для рассеивания семян и опыления, поскольку тогда ещё не было способных это делать насекомых.
Одновременно с насекомыми и животными развивались преобладающие сегодня цветковые (покрытосеменные) растения, поэтому часто опыляются ими.
Простейшие водоросли были древнейшими из известных растений.
Это одноклеточные организмы, всё|все функции которых выполняла единственная лишённая ядра|ядра клетка. Крайне примитивными были эти сине–зелёные водоросли, и только около 1,5 млрд. лет назад появились имеющие клеточное ядро водоросли.
Многоклеточные организмы возникли со временем. Возможно, они сходные с морскими водорослями и имеющие в разных частях растения органы|органы размножения.
Около 590 млн. лет назад, в кембрийский период, прочно обосновались на Земле многие формы жизни. К этому периоду относятся более 900 видов – и это те растения, которые сохранились и были обнаружены сотни миллионов лет спустя.
Читайте также: