
Для преодоления стерильности при размножении отдаленных гибридов у растений используют
Обновлено: 08.07.2024
Методы селекции растений
1. Какая разница с генетической точки зрения между самоопылением и перекрестным опылением?
2. Что такое полиплоидия?
3. Почему большинство культурных растений размножают вегетативно?
Центры происхождения культурных растений.
Основой успеха селекционной работы в значительной степени является генетическое разнообразие исходного материала. В своей работе селекционеры стараются использовать все многообразие диких и культурных растений.
На необходимость использовать в селекции растений все видовое многообразие флоры нашей планеты указывал еще академик Николай Иванович Вавилов, выдающийся генетик и селекционер. Под его руководством были организованы научные экспедиции в разные регионы Земли для сбора образцов культурных растений, их диких предков и сородичей. В ходе экспедиций было собрано более 160 тыс. образцов разных видов и сортов растений.
В настоящее время эта уникальная коллекция хранится во Всесоюзном институте растениеводства и используется селекционерами в их практической работе. Так, известный сорт озимой пшеницы Безостая-1 был получен в результате гибридизации аргентинских пшениц из коллекции Н. И. Вавилова с отечественными сортами.

Работа по созданию семенных коллекций культурных и диких растений продолжается и в наше время. Сейчас коллекция, начало которой положил Н. И. Вавилов, включает более 320 тыс. образцов.
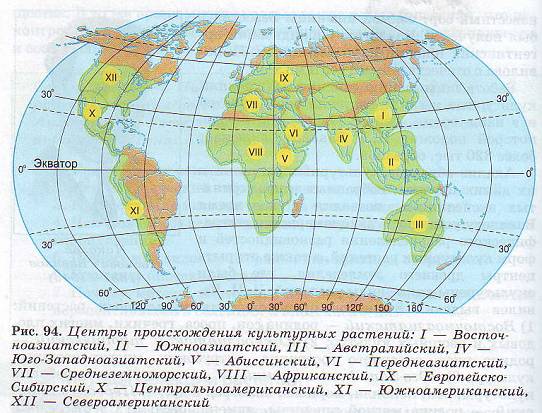
Анализ образцов культурных растений и их диких предков, собранных в предпринятых экспедициях, позволил в свое время Вавилову установить закономерности географического распределения разновидностей и форм культурных растений, а также открыть центры древнего земледелия, где были окультурены дикие виды растений. Н. И. Вавилов выделил 8 центров происхождения культурных растений: 1) Восточноазиатский — родина сои, проса, гречихи, многих плодовых и овощных культур; 2) Южноазиатский тропический — родина риса, сахарного тростника, цитрусовых, многих овощных культур; 3) Юго-Западноазиатский — пшеница, рожь, бобовые культуры, лен, конопля, морковь, виноград и др.; 4) Переднеазиатский — родина мягкой пшеницы, ячменя, овса; 5) Среднеземноморский — родина капусты, свеклы, маслин; б) Абиссинский — родина твердой пшеницы, сорго, бананов, кофе; 7) Центральноамериканский — родина кукурузы, какао, тыквы, табака, хлопчатника; 8) Южноамериканский — родина картофеля, ананаса, хинного дерева.
Дальнейшие исследования ученых привели к установлению еще четырех центров; Австралийского, Африканского, Европейско-Сибирского и Североамериканского (рис. 94).
Закон гомологических рядов наследственной изменчивости.
На примере злаков Н. И. Вавилов показал, что сходные признаки наблюдаются у разных видов данного семейства. Так, у пшеницы, ячменя, овса и кукурузы бывает белая, красная и черная окраска зерновок, существуют голые и пленчатые зерновки, встречаются колосья с длинными и короткими остями, безостые и с вздутиями вместо остей. В ходе последующих наблюдений было выяснено, что данный закон применим не только для растений, но распространяется на животных и микроорганизмы. Так, альбинизм встречается у всех классов позвоночных животных, короткопалость наблюдается у всех пород крупного рогатого скота, овец и собак.
Основные методы селекции растений.
Биологические особенности растений позволяют в селекционной работе с ними использовать инбридинг, полиплоидию, искусственный мутагенез, отдаленную гибридизацию и другие методы.
Отбор и гибридизация являются основными и традиционными методами селекции растений. Применяя массовый или индивидуальный отбор, селекционер не создает ничего нового, а выделяет растения с полезными качествами, уже имеющиеся в популяции. Этим методом выведены многие сорта, в том числе так называемые сорта народной селекции, например знаменитый по своим качествам сорт яблони Антоновка.
Для создания сортов растений с запрограммированными качествами ведется специальная целенаправленная работа — подбирается исходный материал, проводится гибридизация с последующим отбором.
Используя метод гибридизации с последующим отбором, селекционеры получили ценные высокоурожайные сорта пшеницы, ржи, подсолнечника, овощных, плодовых и других культур.
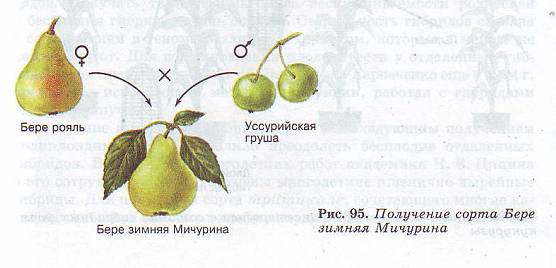
В разработку теории и практики селекции растений большой вклад внес ученый-селекционер Иван Владимирович Мичурин (1855— 1935), Он вывел около 300 новых сортов плодовых растений. В своих работах он широко применял скрещивание географически отдаленных форм. Так, скрещивая французский сорт груши Бере рояль с дикой уссурийской и выращивая сеянцы в условиях средней полосы России, он создал сорт Бере зимняя, сочетающий высокие вкусовые качества плодов с зимостойкостью (рис, 95). Методы, разработанные И. В. Мичуриным, успешно используются селекционерами и в настоящее время.
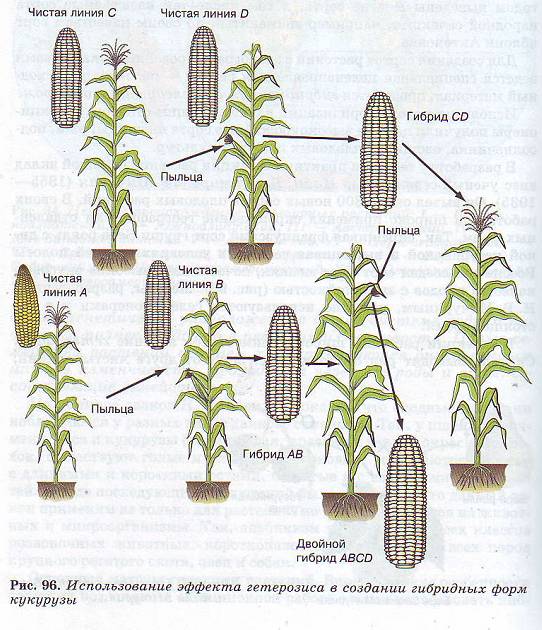
В селекции растений широко применяется явление гетерозиса.
Сначала выводят ряд отличающихся друг от друга чистых линий, а затем производят межлинейное скрещивание.
Выяснив, в каких случаях эффект гетерозиса проявляется наиболее сильно, используют лишь эти линии для получения гибридных семян. Эта методика применяется для получения высоких урожаев кукурузы, огурцов, томатов и других культур (рис. 96).

Полиплоидию (кратное увеличение числа хромосом) издавна использовали при создании сортов пшеницы, овса, картофеля, хлопчатника, плодовых, декоративных и других культур. Полиплоидные растения появлялись в популяциях случайно в результате естественных мутаций. В настоящее время применяют методы искусственного получения полиплоидов, воздействуя на растения разными мутагенами (в основном колхицином), разрушающими веретено деления клетки. Таким образом из диплоидных (2n) можно получить тетраплоидные (4n) формы.
Большинство их неперспективны, но отдельные формы служат ценным материалом для гибридизации и отбора. Полиплоидные растения могут отличаться более крупными размерами, высокой урожайностью и более активным синтезом органических веществ. Использование метода полиплоидии позволило селекционерам получить ценные сорта сахарной свеклы, ржи, гречихи, фасоли и других культур (рис. 97).
Отдаленная гибридизация позволяет в одном организме совместить признаки, характерные для растений разных видов и даже родов. Получать такие формы из-за нескрещиваемости родителей и бесплодия гибридов очень сложно. Стерильность гибридов связана с содержанием в геноме различных хромосом, которые в мейозе не конъюгируют. Для восстановления плодовитости у отдаленных гибридов известный генетик Георгий Дмитриевич Карпеченко еще в 1924 г. предложил использовать метод полиплоидии, работая с гибридами редьки и капусты.
Сочетание отдаленной гибридизации с последующим получением полиплоидных форм позволило преодолеть бесплодие отдаленных гибридов. В результате многолетних работ академика Н. В. Цицина и его сотрудников были получены многолетние пшенично-пырейные гибриды. Для получения сорта тритикале, сочетающего многие качества пшеницы (высокие хлебопекарные качества) и ржи (высокое содержание незаменимой аминокислоты лизина, а также способность расти на бедных песчаных почвах), применялась следующая схема:
Р: пшеница (2n = 42) х рожь (2n = 14)
G : n = 21
F1 2n = 28 (все непарные)
G: Мейоз нарушен, гибрид стерилен, нормальных гамет нет.
Обработка колхицином приводит к удвоению числа хромосом,
F1 :(колхицированное): 2n = 56
G : n = 28
F2, F3, , Fn : 2n= 56 (тритикале)
У таких гибридов в клетках содержится полный диплоидный набор хромосом обоих родителей, поэтому их хромосомы конъюгируют друг с другом и мейоз проходит нормально.
С помощью метода отдаленной гибридизации с последующим получением полиплоидных форм были выведены новые перспективные сорта картофеля, табака и других культур.
Методами отдаленной гибридизации и радиационного мутагенеза созданы перспективные сорта хлопчатника. Химический мутагенез лежал в основе получения многих новых сортов кукурузы, пшеницы, риса, овса, подсолнечника.
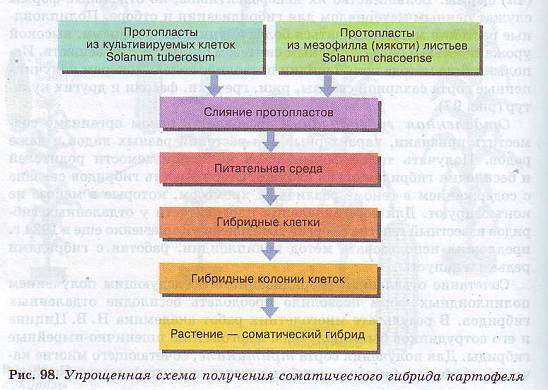
Методы клеточной инженерии.
Селекционеры все шире начинают применять для получения новых сортов растений методы клеточной инженерии. В качестве примера можно привести работу по соматической гибридизации двух видов картофеля: культурного — Solanum tuberosum и дикого Solanum chacoense (рис. 98). Для гибридизации использовались протопласты (греч. protos — первый и греч. plastos — вылепленный, образованный) — клетки, полностью лишенные клеточной стенки (оболочки) и имеющие только клеточную мембрану, которая ограничивает цитоплазму с различными органоидами.
Полученный соматический гибрид в сравнении с родительскими формами имел промежуточные характеристики по форме листа, величине клубней, но отличался большей мощностью куста и высотой стеблей, благодаря чему и был включен в дальнейшую практическую селекционную работу (рис. 99).

Метод вегетативного размножения культурой тканей широко применяется в селекции для быстрого размножения новых перспективных сортов растений.
В различных регионах нашей страны созданы научно-исследовательские институты и селекционные станции, которые проводят работы по выведению и районированию новых сортов растений. Эта работа играет важнейшую роль в повышении урожайности сельскохозяйственных культур и обеспечении населения продовольствием.
Центры происхождения культурных растений. Закон гомологических рядов наследственной изменчивости.
1. Какие методы применяются в селекции растений?
2. Какое значение для селекции имеет открытие закона гомологических рядов наследственной изменчивости?
3. Почему межлинейные гибриды сохраняют ценные признаки при вегетативном размножении и теряют их при семенном?
4. Почему селекционеры стремятся Получить растения-полиплоиды?
5. Какая методика позволяет преодолеть стерильность межвидовых (межродовых) гибридов?
Онлайн библиотека с учениками и книгами, плани-конспекти уроковс Биологии 10 класса, книги и учебники согласно календарного плана планирование Биологии 10 класса
Если у вас есть исправления или предложения к данному уроку, напишите нам.
Если вы хотите увидеть другие корректировки и пожелания к урокам, смотрите здесь - Образовательный форум.

Биология ЕГЭ 100БАЛЛОВ запись закреплена
Для преодоления стерильности при половом размн-ии гибридов растений используют
Для преодоления стерильности гибридов первого поколения применяют различные методы, из которых можно выделить два главных: 1) возвратные скрещивания, 2) удвоение числа хромосом у гибридных растений для получения аллополиплоида.
Применение возвратных скрещиваний основано на том, что женские гаметы гибрида обычно обладают большей жизнеспособностью, чем мужские. Использование для опыления гибрида нормальной пыльцы одной из родительских форм позволяет получить семена для дальнейшей работы. С этой же целью гибриды первого поколения можно опылять пыльцой третьего родственного вида, например: (рожь x пшеница) x пырей; (рожь x пырей) x пшеница; (пшеница x пырей) x рожь.
Наиболее надежным методом преодоления стерильности межвидовых и межродовых гибридов F1 является удвоение у них числа хромосом. Поскольку у полученных таким путем амфидиплоидов каждый тип хромосом представлен парой, то мейоз протекает сравнительно нормально с образованием жизнеспособных гамет, содержащих по одному геному скрещиваемых видов. Иногда плодовитые амфидиплоиды удается обнаружить в небольшом количестве среди большой массы обычно бесплодных гибридных растений. Разумеется, в редких случаях гаметы могут формироваться и у амфигаплоидных гибридных растений F1.
Преодолению несовместимости разных видов и стерильности их гибридов могут способствовать и некоторые другие приемы, например создание благоприятных условий во время цветения растений, применение физиологически активных веществ, химических мутагенов и других факторов.
Количество завязывающихся семян на гибридных растениях F1 зависит также от общего числа цветков, которое можно увеличить созданием наиболее благоприятных условий для формирования генеративных органов, а также путем вегетативного размножения растений. Разработаны достаточно эффективные способы клонирования разных видов растений, в том числе злаковых, например риса и др.
а) Отдаленная гибридизация - скрещивание особей, принадлежащих к разным видам, родам, с целью объединения у гибридов ценных наследственных признаков.
Рожь х пшеница - тритикале;
Кобыла х осел - мул.
б) Методику преодоления стерильности межвидовых гибридов у растений разработал в 1924г. Г.Д. Карпеченко на примере скрещивания редьки и капусты.
5.Растения как объекты селекции:
1) характерно как половое, так и бесполое размножение (вегетативное)
2) большое количество потомков;
3) неприхотливы к условиям среды;
4) незначительные экономические издержки;
6.Методы селекции растений.
1) Использование различных форм искусственного отбора: индивидуальный и массовый;
2) Использование различных форм гибридизации: родственное, неродственное, межвидовое скрещивание;
3) полиплоидия как метод преодоления стерильности растительных межвидовых гибридов. Возникновение полипоидов у растений в результате удвоения хромосом, которое не сопровождается делением клетки; слиянием соматических клеток; образованием гамет с не редуцированным числом хромосом. Полиплоидные растения характеризуются интенсивным ростом, большими размерами и массой плодов и семян, повышенной устойчивостью к неблагоприятным факторам и болезней;
4) прививки - особый способ искусственного объединения различных частей растений. Прививка не является настоящей гибридизацией, ибо приводит лишь к не наследственным изменениям фенотипа.
7.Особенности селекции животных.
Животные как объекты селекции:
1) характерно только половое размножение;
2) немногочисленное потомство;
3) каждый объект является значительной селекционной ценностью;
4) сложные взаимодействия с окружающей средой через развитую нервную систему;
5) значительные экономические затраты на содержание.
8.Методы селекции животных.
Те, что и для растений. Отличия заключаются в том, что в селекции животных не применяют массовый отбор, учитывая коррелятивную изменчивость.
В селекции животных используют искусственное оплодотворение получения ценных пород КРС в искусственных условиях.
9.Особенности селекции микроорганизмов.
Микроорганизм как объект селекции:
1) не имеют полового процесса (прокариоты);
2) имеют гаплоидный набор хромосом или кольцевую молекулу ДНК, что позволяет мутациям проявиться уже в первом поколении потомков;
3) быстрые темпы размножения позволяют получить большое количество клеток-потомков.
21. Методы преодоления нескрещиваемости отдалённых видов и родов. Преодоление нескрещиваемости генетически далёких видов. В основе всех причин нескрещиваемости лежит генетическая дифференциация видов, их генетическая изоляция, которые выражаются в несовместимости генотипов видов. Существуют генетическое, физиологическое и структурное несоответствия гамет отдалённых форм. Степень скрещиваемости в значительной мере определяется филогенетическим родством скрещиваемых форм. В связи с этим выделяют две основные причины трудной скрещиваемости при отдалённой гибридизации:
а) Препятствия к опылению и оплодотворению: несовпадение циклов размножения (несовпадение циклов развития гамет), несовместимость пыльцевых трубок с тканью пестика, генетическая несовместимость ядер, несущих разные геномы (несовместимость яйцеклеток и спермиев), физиологическая несовместимость ядра и цитоплазмы.
б) Нежизнеспособность (бесплодие) или малая жизнеспособность (низкая плодовитость) гибридной зиготы (зародыша).
И.В. Мичуриным и другими исследователями были предложены следующие методы преодоления нескрещиваемости:
– Предварительная прививка. Прививка одного растения на другое изменяет химический состав тканей, осмотическое давление в клетках и т.д. Это увеличивает вероятность прорастания чужих пыльцевых трубок в пестике материнского растения. Например, пшенично-элимусные гибриды были получены путём предварительной пересадки зародыша пшеницы на эндосперм элимуса.
– Метод посредника (метод мостов) состоит в том, что нескрещиваемость двух видов преодолевается с помощью третьего вида. Если виды А и В не скрещиваются между собой, то вид А скрещивают с близким видом С, а полученный гибрид – с видом В. В результате в гибриде могут быть совмещены хромосомы и признаки трёх видов. Данный метод нашёл применение в селекции картофеля, пшеницы, овса и других культур. И.В. Мичурин при скрещивании персика с бобовником использовал в качестве посредника дальневосточный персик Давида Диплоидные виды овса A. strigosa и A. pilosa не скрещиваются между собой, но успешно гибридизируются с A. longiglumis. Гибриды между данными диплоидными видами и A. Longiglumis легко скрещиваются между собой.
– Опыление смесью пыльцы разных видов также повышает скрещиваемость за счёт того, что пыльца, имеющая разный генотип, может взаимно стимулировать рост составных частей её, создавая в пестике условия, благоприятные для прорастания разной пыльцы. В данном случае используют смесь пыльцы различных биотипов. Подобные явления происходят при использовании таких приёмов, как повторное опыление и доопыление материнской пыльцой.
– Использование разных биотипов. При получении гибридов Raphanobrassica завязываемость семян при скрещивании редьки масличной с сортом кормовой капусты Синий гигант была в 7–10 раз выше, чем от скрещивания с сортом капусты Мозговая зелёная вологодская.
– Изменение уровня плоидности у родительских форм. Перевод скрещиваемых видов на тетраплоидный уровень способствует получению гибридных семян, что свидетельствует об изменении селективности оплодотворения.
– Предварительное воздействие на скрещиваемые растения химическими и физическими факторами – радиацией, химическими мутагенами, биологически активными веществами, стрессовыми условиями выращивания. Применение стероидных глюкозидов при межвидовой гибридизации томатов приводило к повышению завязываемости гибридных семян. Обработка пестиков стимуляторами роста и другими химическими веществами позволяет изменить активность ферментов, рН среды в пестике в период оплодотворения. Например, в СИММИТе (Мексика) для преодоления генетической несовместимости исходных родительских форм применяют обработку родительских растений в период формирования у них генеративных органов 0,1%-ным раствором эпсилон-амино-капроновой кислоты (опрыскиванием или инъекцией). Таким путём удалось скрестить пшеницу с ячменём.
– Перенос скрещиваемых форм в другие экологические условия. На скрещиваемость видов в сильной степени влияет различное физиологическое состояние генеративных органов растений, которое зависит от факторов внешней среды. Преодоления фенотипической несовместимости видов иногда можно добиться путём переноса скрещиваемых растений в иные экологические условия. Например, в Закавказье скрещивание между разными видами пшеницы происходит значительно легче, чем в другой зоне.
– Использование метода гибридизации соматических клеток (парасексуальная гибридизация) путём слияния изолированных протопластов. Таким путём можно получать, например, ЦМС аналоги культурных растений, без применения насыщающих скрещиваний. Гены цитоплазмы кодируют ряд признаков, представляющих практический интерес (ЦМС, скорость фотосинтеза, устойчивость к патогенам и абиотическим факторам). С помощью соматической гибридизации возможно получение цибридов, совмещающих пластомы двух видов. Путём соматической гибридизации получены гибриды картофеля с томатом, капусты с турнепсом, культурного картофеля с диким и др. Кроме рассмотренных методов для преодоления нескрещиваемости в селекции используют культивирование на питательной среде вычлененных семяпочек, проводят опыление на ранних этапах развития рыльца, укорачивают столбик или удаляют рыльце перед опылением с заменой его кусочком питательной смеси и т. д.
22. Методы преодоления невсхожести гибридных семян и стерильности отдалённых гибридов. Преодоление неспособности гибридных семян к прорастанию осуществляется путём применения метода культуры зародышей и тканей в стерильных условиях. Эндосперм является как бы промежуточным звеном между зародышем и материнским растением. Он несёт ответственность за отклонения в развитии гибридных зёрен. Поэтому различные нарушения в развитии эндосперма (замедленное формирование клеток, образование гигантских ядер, нарушения в образовании хлорофиллоносного слоя и клеток перикарпия и др.) отрицательно влияют на гибридный зародыш. Метод культуры зародышей широко используют, например, при получении пшенично-элимусных гибридов. Во многих случаях прорастанию зародышей способствуют применение биологически активных веществ и создание определённых условий для прохождения длительного покоя (приём часто применяют для семян с твёрдой оболочкой). Для определения степени жизнеспособности гибридных семян индийскими селекционерами разработан рентгеноскопический метод анализа, позволяющий отделять здоровые семена от недоразвитых и пустых.
3) преодоление стерильности (бесплодия) гибридов первого поколения. Как правило, чем дальше в филогенетическом отношении отстоят друг от друга скрещиваемые формы, тем сильнее выражена стерильность их гибридов. Стерильность гибридов F1 вызывается следующими причинами: а) недоразвитием генеративных органов (чаще всего пыльников); б) абортивностью зародыша из-за несоответствия зиготы и эндосперма; в) нарушениями в мейозе и митозе, приводящими к появлению нежизнеспособной пыльцы и аномальных яйцеклеток. Нарушения в мейозе и митозе вызываются:
– Различиями в структуре, числе хромосом и генных наборов у скрещиваемых форм, т.е. несовместимостью геномов, хромосом и отдельных генов, следствием чего является нарушение конъюгации хромосом в мейозе (асинапсис, десинапсис, образование унивалентов). Хромосомная стерильность наблюдается, например, у пшенично-элимусных, пшенично-ржаных, ржано-пырейных и других гибридов.
– Несовместимостью ядра и цитоплазмы у родительских форм, что приводит к нарушению митотического деления клеток в процессе образования генеративных органов, что проявляется в стерильности пыльцы гибридных растений. Действие отдельных генов препятствует развитию мужских и женских органов цветка. Проростки могут гибнуть на ранней стадии их развития.
– геномной аллоциклией, которая вызывается различиями в темпах деления хромосом исходных видов. Так, у мягкой пшеницы темп деления хромосом 24 ч, твёрдой – 30 ч, ржи – 51, тритикале – 34–37 ч. Поэтому у тритикале часто происходит потеря хромосом ржи. На проявление геномной аллоциклии влияет цитоплазма материнского растения и условия выращивания. Разработаны следующие методы преодоления стерильности (восстановления плодовитости) гибридов F1:
– Метод последовательных насыщающих (возвратных) скрещиваний, который позволяет восстановить хромосомный набор одного из родителей, но с новым сочетанием генов. Применение возвратных скрещиваний основано на том, что женские гаметы гибрида обычно обладают большей жизнеспособностью, чем мужские. Для опыления гибрида используют нормальную пыльцу одной из родительских форм. Гибриды первого поколения можно также опылять пыльцой третьего родственного вида, например: (рожь х пшеница) х пырей; (рожь х пырей) х пшеница; (пшеница х пырей) х рожь.
– Удвоение числа хромосом у гибридов F1 с помощью колхицина, или получение амфидиплоидов. Амфидиплоиды (аллотетраплоиды) – полиплоиды, возникающие в результате соединения и последующего удвоения хромосомных наборов двух разных видов или родов (ААВВ). Диплоид АВ от скрещивания АА х ВВ отличается полной стерильностью (например, пшенично-ржаные амфидиплоиды – тритикале). При удвоении числа хромосом каждая хромосома имеет гомологичную хромосому, в результате чего конъюгация протекает нормально и плодовитость гибридов восстанавливается. Увеличить количество завязавшихся семян на гибридном растении можно путём создания благоприятных условий для формирования генеративных органов (во время цветения), с помощью вегетативного размножения, например клонирования злаков и других растений, путём обработки физиологически активными веществами и др.
23. Пути передачи признаков при отдалённой гибридизации. Возможность передачи генов от одного вида другому зависит от филогенетической близости скрещиваемых форм. Выделяют пять основных методов межвидовой передачи признаков:
1. Генетическая рекомбинация на основе кроссинговера. При большой геномной близости скрещиваемых видов работа с гибридными поколениями ведётся в основном также, как и при межсортовой гибридизации, и опирается на генетическую рекомбинацию. Селекционер в большинстве случаев ставит задачу передать культурному растению от второго вида лишь отдельный ген или признак, например устойчивость к какой-нибудь болезни. Наиболее эффективный метод для достижения этой цели – возвратные скрещивания. В итоге получают интрогрессивную форму исходного сорта, включающую от второго вида лишь единичные признаки. Например, Н.В. Цицин вывел ценные сорта пшеницы от скрещивания с пыреем. М.Ф. Терновский путём передачи культурному табаку генов устойчивости от вида N. glutinosa L. получил иммунные к табачной мозаике и мучнистой росе сорта: Американ 187-С, Дюбек 566, Иммунный 580 и др. При нарушении конъюгации хромосом в мейозе у гибридов передать отдельные признаки от одного вида другому трудно.
2. Синтез амфидиплоидов. Амфидиплоиды получают двумя путями: 1) проводят гибридизацию с последующим удвоением числа хромосом у гибрида, 2) скрещивают предварительно полученные автотетраплоиды исходных видов. Амфидиплоиды могут быть использованы главным образом как исходный материал для селекции. Известны только единичные случаи, когда амфидиплоиды непосредственно используются для практических целей. Например, большое число работ выполнено по пшенично-ржаным амфидиплоидам – тритикале.
3. Добавление и замещение хромосом. Если передачу отдельных генов путём генетической рекомбинации трудно осуществить, то в генотип улучшаемой культуры добавляют отдельную пару хромосом с нужными селекционеру генами от вида-донора. Методика получения линий с добавленными хромосомами описана О’Мара (O’Mara, 1940) на примере получения линий пшеницы с добавленными хромосомами ржи. Линии с добавочными хромосомами получают, осуществляя последовательно следующие процедуры: 1) скрещивают растения разных видов; 2) удваивают число хромосом у гибридов F1; 3) проводят возвратные скрещивания полученного амфидиплоида с рекуррентным родителем (улучшаемый сорт); 4) проводят отбор линий с добавочными хромосомами.
К настоящему времени имеются дополненные линии мягкой и твёрдой пшеницы с парой хромосом от следующих видов: Secale cereale, Aegilops caudata, Ae. umbellulata, Ae. comosa, Agropyron elongatum, Agr. intermedium, Haynaldia villosa. Серия дополненных линий получена также у овса (Avena sativa). В каждой линии к хромосомам овса добавлено по одной хромосоме Avena hirtula. Линии с замещёнными хромосомами получают различными путями, обычно методом возвратных скрещиваний, используя в качестве родителя-донора линию с добавленной парой хромосом от другого вида, а в качестве рекуррентного родителя – линии моносомика или нуллисомика. В настоящее время созданы линии, в которых пара хромосом пшеницы замещена парой хромосом видов Secale cereale, Aegilops comosa, Ae. ventricosa, Agropyron intermedium, Agr. elongatum. У табака были использованы хромосомы Nicotiana glutinosa и N. plumbaginifolia для замещения хромосом N. tabacum с целью передать этому виду устойчивость к табачной мозаике.
Возможны замещения не только пары хромосом, но и отдельных хромосом. Например, Р. Райли передал пшенице мягкой от эгилопса одну хромосому, контролирующую устойчивость к жёлтой ржавчине.
Успех использования дополненных и замещённых линий, также как и амфидиплоидов, зависит от их стабильности и озернённости. К сожалению, все полученные дополненные линии цитологически не стабильны, по-видимому, из-за нарушения сбалансированности функций генов. Наблюдается утеря пары добавленных хромосом и возврат к 42-хромосомным пшеницам. Замещение отдельных хромосом даёт лучший эффект, чем добавление чужой хромосомы, поскольку генотип в данном случае более сбалансирован, однако эти линии часто маложизнеспособны, т. к. чужая хромосома может не компенсировать отсутствующую хромосому.
1. Популяция исходного материала должна быть достаточно многочисленной (от 10 до 20 тыс. особей), чтобы обеспечить генетическое разнообразие. При увеличении размера популяции повышается вероятность отбора ценных генотипов.
2. Отбор элитных растений из исходного материала проводят на основе запланированных критериев отбора в течение всей вегетации. Окончательно элитные растения отбирают по сумме всех критериев отбора после их проявления.
3. Число элитных растений, которое следует отбирать, определяется запланированной интенсивностью отбора. Хороший результат (сдвига при отборе) получают, если отбирают около 10% особей исходной популяции или меньше.
4. Надёжность проверки результата отбора на втором этапе (испытание потомств элитных растений) зависит от точности полевого опыта. Первое потомство испытывают без повторности (так как мало семян), через 10–40 делянок высевают стандартный сорт (районированный в данной местности сорт, широко возделываемый в производстве) или исходную популяцию. Результаты отбора при испытании первого потомства проверяют только по моногенно обусловленным признакам (устойчивость к болезням) или признакам с высокой наследуемостью. Высокий коэффициент наследуемости у зерновых культур имеют такие признаки, как высота растений, длина колоса, масса одного зерна. Потомства с явно низкой урожайностью или восприимчивые к болезням бракуются. На основании испытания первого потомства элитных растений опять отбирают около 10% наилучших линий или семей.
5. Оценку второго поколения потомств проводят в точном полевом опыте с повторностями. Теперь результат отбора можно проверить по урожайности и другим признакам с низкой и средней наследуемостью. Из этих потомств отбирают опять около 10% наиболее продуктивных.
6. Площадь делянок и число повторностей в каждом последующем испытании увеличивается, а число потомств вследствие отбора уменьшается. Испытание прекращают, когда можно указать потомства, наилучшим образом отвечающие задачам селекции. Как правило, проводят три испытания потомств, из которых два последних – в местах будущего распространения создаваемых сортов.
25 планирование отбора , селекционный дифференциал , р-ция на отбор , коэффициент наследуемости.
При условии промежуточного наследования, частичного или полного доминирования эффект отбора количественным признакам зависит от его интенсивности и силы модифицирующего влияния внешней среды.
Читайте также: