
Двойное оплодотворение у растений реферат
Обновлено: 05.10.2024
Двойное оплодотворение, половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений — лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго — с полярными ядрами или со вторичным ядром зародышевого мешка.
Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки — эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой.
Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой — проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое — ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое — мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы.
При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден.
Следствие двойного оплодотворения — ксении — проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных — дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез, а также опыление.
, половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon ) и рябчике (Fritillaria orientalis ). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки - эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие двойного оплодотворения - ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез , а также опыление .
Микроспорогенез протекает в пыльниках тычинок. При этом диплоидные клетки образовательной ткани пыльника в результате мейоза превращаются в 4 гаплоидные микроспоры. Через некоторое время микроспора приступает к митотическому делению и преобразуется в мужской гаметофит – пыльцевое зерно .
Мегаспорогенез осуществляется в семязачатке . Из материнской клетки нуцеллуса в результате мейоза образуются 4 мегаспоры, из которых в результате остается только одна. Эта мегаспора сильно разрастается и оттесняет ткани нуцеллуса к интегументам, формируя зародышевый мешок . Ядро зародышевого мешка делится 3 раза митозом. После первого деления два дочерних ядра расходятся к разным полюсам: халазальному и микропилярному , и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На микропилярном полюсе находятся две синергиды и одна более крупная клетка – яйцеклетка . На халазальном полюсе располагаются антиподы . Таким образом, зрелый зародышевыый мешок содержит 7 клеток.
Опыление заключается в переносе пыльцы с тычинок на рыльце пестика.
Оплодотворение . Пыльцевые зерна, тем или иным образом попавшие на рыльце, - прорастают. Прорастание пыльцы начинается с разбухания зерна и образования пыльцевой трубки из вегетативной клетки. Пыльцевая трубка прорывает оболочку в более тонком ее месте – так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика. По мере роста пыльцевой трубки в нее переходит ядро вегетативной клетки и генеративной, которая делится и образует два спермия. Через микропиле семязачатка пыльцевая трубка проникает в зародышевый мешок, там она разрывается, и ее содержимое изливается внутрь. Один из спермиев сливается с яйцеклеткой, образуя зиготу, которая затем дает начало зародышу семени. Второй спермий сливается с центральным ядром, что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Таким образом, эндосперм у покрытосеменных триплоидный и вторичный , т.к. образуется после оплодотворения.
Весь этот процесс получил название двойного оплодотворения . Он был впервые описан русским ученым С.Г.Навашиным. (1898 г.).
У голосеменных в оплодотворении участвует один спермий и эндосперм возникает до оплодотворения в результате деления мегаспоры, т.е. он гаплоидный и первичный .
Антиподы и синергиды растворяются после оплодотворения, а интегументы преобразуются в семенную кожуру.
Апомиксис – развитие зародыша из неоплодотворенной клетки. Формы апомиксиса зависят от того, из каких частей семязачатка развивается зародыш. При партеногенезе (партенос- девственный) зародыш возникает из неоплодотворенной яйцеклетки. Если зародыш развивается из какой-либо другой клетки гаметофита (антиподы, синергиды), то такой процесс называется апогамия . В случае образования зародыша из клеток нуцеллуса, интегументов, не входящих в гаметофит, говорят об апоспории . Апомиксис распространен среди эволюционно развитых групп растений. За счет апомиксиса растения не зависят от опыляющих агентов

ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ половой процесс у покрытосеменных растений, при к-ром оплодотворяются как яйцеклетка, так и центр. клетка зародышевого мешка. Д. о. открыл рус. учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis) . В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центр. клетки - эндосперм. В зародышевых мешках с трёхклеточным лицевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синер-гиды и вегетативного ядра пыльцевой трубки) ; вторая синергида впоследствии отмирает. Далее оба спермия вместе с изменённой цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центр. клеткой. Затем спермин разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним,а иногда и с обоими полярными ядрами. Спермин теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермин в виде неизменённых клеток наблюдаются и в
зародышевом мешке.
Двойное оплодотворение; 1 - у рябчика: один из спермиев (а) в контакте с ядром яйцеклетки, второй (б) - с одним из полярных ядер (второе полярное ядро не изображено); 2 - у подсолнечника; а - пыльцевая трубка; 6 - синергиды (одна из них повреждена пыльцевой трубкой); в - яйцеклетка; г - спермий в контакте с ядром яйцеклетки; д - центральная клетка; е - второй спермий в контакте со вторичным ядром зародышевого мешка.
При Д. о. ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние к-рых могут вырьировать. У скерды и нек-рых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у мн. растений они удлинённые, иногда извитые, б. или м. хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Ге-расимова-Навашина) различать два типа Д. о.: премитотическое -ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе) ; постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце к-рой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоид-но. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центр. клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплои-ден. Следствие Д. о. - Ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает неск. пыльцевых трубок, спермий первой из них участвуют в Д. о., спермии остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Рядом точных эмбриологических исследований на различных представителях покрытосеменных (лилейных, лютиковых, сложноцветных) С. Г. Навашин убедительно показал, что эндосперм, как и зародыш, является продуктом полового процесса. Это необычное, свойственное лишь покрытосеменным растениям, явление он назвал двойным оплодотворением. О своем открытии он сообщил в августе 1898 г. на проходившем в Киеве Х съезде русских естествоиспытателей и врачей, а в ноябре того же года опубликовал на эту тему небольшую статью в "Известиях Петербургской Академии Наук".
Мысль о существовании двойного оплодотворения возникла у С. Г. Навашина еще в 1895 г. в процессе работы над изучением халазогамии у грецкого ореха. Окончательное же подтверждение и оформление в стройную теорию эта мысль получила при исследовании оплодотворения у лилии.
Позднее С. Г. Навашин описал двойное оплодотворение и у других цветковых растений, систематически далеко отстоящих друг от друга - у представителей лютиковых, сложноцветных, орехоцветных, доказав тем самым общность этого явления для всех покрытосеменных.
Двойное оплодотворение - отличительный признак, отделяющий покрытосеменных от голосеменных.
Открытие С. Г. Навашиным двойного оплодотворения у покрытосеменных сыграло важную роль в науке.
Явление двойного оплодотворения внесло ясность не только в вопрос происхождения эндосперма, но и разъяснило загадочность такого явления, как ксения у кукурузы. Работы Навашина по оплодотворению покрытосеменных растений были встречены ботаниками всего мира с огромным интересом.
Дискуссии о сущности двойного оплодотворения продолжались и в начале ХХ в. В ходе этих дискуссий, а главное, благодаря новым исследованиям, выводы Навашина получили полное подтверждение и дальнейшее развитие.
Двойное оплодотворение Цветковых растений – это процесс, в результате которого один из спермиев, идущих по пыльцевой трубке, сливается с яйцеклеткой, а другой – с центральной клеткой. В результате слияния спермия с яйцеклеткой образуется диплоидный зародыш - будущий спорофит. Центральная клетка уже имеет диплоидный набор хромосом, так как она образовалась после слияния двух из первоначальных восьми клеток зародышевого мешка. После слияния со спермием образуется триплоидная клетка, которая дает начало эндосперму.
У Голосемянных один из спермиев, идущих по пыльцевой трубке, погибает, а другой оплодотворяет яйцеклетку с образованием зародыша. Эндосперм же в данном случае представляет собой гаметофит и гомологичен заростку Папоротникообразных. Следовательно, эндосперм Голосемянных гаплоиден.
Чтобы понять, какие преимущества имеет триплоидный эндосперм над гаплоидным, разбересначала, что такое эндосперм и какие у него функции. Эндосперм – это запасающая ткань семени растений, в которой откладываются питательные вещества для дальнейшего развития и прорастания семени. Он представлен тонкостенными крупными клетками часто неправильной формы, заполненными крахмалом и белками. Функции эндосперма неразнообразны. Первая и главная функция эндосперма – это питательная. Возможно также участие в прорастании семени.
Первое и очевидное преимущество триплоидного эндосперма над гаплоидным в проявлении мутаций. Если в генах гаплоидного эндосперма происходит мутация, то она обязательно проявится. А это может привести к недостаточному выполнению функций или даже к редукции эндосперма. В результате семя не разовьется вообще, или растение будет слаборазвитым. В триплоидном же эндосперме рецессивная мутация проявляться не будет. Следовательно, в триплоидном эндосперме по сравнению с гаплоидным увеличивается вероятность нормального развития семени даже при наличии мутаций. Кроме того, если хромосомный набор в клетке 3n, то интенсивность транскрипции и, соответственно, трансляции и биосинтеза белка в клетке в три раза выше, чем в таковой же с гаплоидным набором хромосом. С интенсивностью транскрипции косвенно связано и участие эндосперма в прорастании семени. Как известно, прорастание инициируется множеством факторов таких, как кратковременное воздействие низких температур, для некоторых семян воздействие света и т. д. Но непосредственно перед прорастанием клетки начинают интенсивно поглощать воду, клетки зародыша набухают и растягиваются. Для процессов прорастания растению нужно много энергии и необходимы ферменты, катализирующие расщепление крахмала и белков. Под воздействием воды в клетках зародыша активируются гиббереллины, они транспортируются в тонкий верхний слой живых клеток эндосперма (алейроновый слой), где сразу же начинают синтезироваться амилазы и протеазы. В соответствии с выше сказанным предположением о скорости транскрипции в триплоидных клетках нужное количество ферментов выработается быстрее, чем в гаплоидных, и процесс прорастания будет идти интенсивнее.
Рассмотрим, есть ли предпосылки усложнения процесса оплодотворения и появления тройного оплодотворения. Раз оплодотворение тройное, значит, спермиев должно быть три. Один должен сливаться с яйцеклеткой с образованием зародыша. Слияние двух остальных спермиев с разными структурами просто не целесообразно, так как семени нужно образование двух структур в ходе оплодотворения: зародыша и запасающей ткани (в данном случае эндосперма). Тогда получается, что оставшиеся 2 спермия должны оба слиться с центральной клеткой, и получится тетраплоидный эндосперм. Механизм этого сложно представить. Либо два спермия должны по очереди слиться с центральной клеткой, что маловероятно, потому что после оплодотворения одним спермием сразу включаются механизмы блокирования оплодотворения другим спермием. Либо 2 спермия должны слиться заранее в пыльцевой трубке, что в общем-то тоже трудно себе представить. Кроме того, в результате такого оплодотворения образуется диплоидный зародыш и тетраплоидный эндосперм, а это равносильно гаплоидному зародышу и диплоидному эндосперму. При дальнейшем увеличении плоидности эндосперма будут возникать проблемы с расхождением хромосом, сильно возрастет частота мутаций, возникнет проблема размещения хромосом в ядре. Следовательно, предпосылок к усложнению процесса оплодотворения нет.
Похожие страницы:
Генетика. Учебно-методическое пособие для самостоятельных занятий
. Изложить генетическое и биологическое значение двойного оплодотворения. Запомнить хромосомный набор основных сельскохозяйственных . . Женский гаметофит. Макроспорогенез и макрогаметогенез. Двойное оплодотворение у растений, его генетический и биологический .
Отдел покрытосеменные цветковые
. Навашиным двойного оплодотворения у покрытосеменных сыграло важную роль в науке. Явление двойного оплодотворения внесло . Навашиным, получил в науке название двойного оплодотворения. Дискуссии о сущности двойного оплодотворения продолжались и в начале ХХ в. .
Теоретические вопросы генной инженерии
. особый генеративный орган – цветок. Имеется двойное оплодотворение и триплоидный эндосперм Имеют 1-2 семядоли . формируется эндосперм. Открыл двойное оплодотворение Навашин. Биологический смысл двойного оплодотворения Первичный эндосперм голосеменных .
Растения, грибы, бактерии и лишайники
. насекомыми. Семя Семя формируется в результате двойного оплодотворения из семенного зачатка. Цветковые растения . развиваются органы спороношения — тычинки и пестики; оплодотворение двойное; перенос пыльцы (опыление) осуществляется различными .
Бесполое и половое размножение
. самооплодотворению и благоприятствующие перекрестному оплодотворению. Например, у многих простейших . У цветковых растений происходит двойное оплодотворение. При опылении пыльцевое зерно . запас питательных веществ. Двойное оплодотворение у цветковых растений
Функция "чтения" служит для ознакомления с работой. Разметка, таблицы и картинки документа могут отображаться неверно или не в полном объёме!
Двойное оплодотворение, половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки - эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие двойного оплодотворения - ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез, а также опыление.
Микроспорогенез
Похожие работы
2014-2022 © "РефератКо"
электронная библиотека студента.
Банк рефератов, все рефераты скачать бесплатно и без регистрации.
"РефератКо" - электронная библиотека учебных, творческих и аналитических работ, банк рефератов. Огромная база из более 766 000 рефератов. Кроме рефератов есть ещё много дипломов, курсовых работ, лекций, методичек, резюме, сочинений, учебников и много других учебных и научных работ. На сайте не нужна регистрация или плата за доступ. Всё содержимое библиотеки полностью доступно для скачивания анонимному пользователю
В 7 классе на уроках ботаники школьники изучают половое размножение покрытосеменных растений. Это помогает детям понять, какой сложный путь прошла флора нашей планеты во время эволюции. Двойное оплодотворение — это уникальное явление. Оно не встречается ни у одного из живых организмов, кроме цветковых растений. Если описывать этот процесс кратко, то можно сказать, что он заключается в оплодотворении не только яйцеклетки, но и центральной клетки зародышевого мешка.

Виды размножения высших растений
Двойное оплодотворение у цветковых растений было впервые исследовано в 1898 году. Его открыл русский ученый Сергей Гаврилович Навашин при исследовании размножения лилии кудреватой и рябчика восточного. Это открытие внесло серьезный вклад в биологию. До этого ученые неоднократно исследовали высшие растения, но никто не обратил внимания на двойное оплодотворение.
Покрытосеменные растения могут размножаться двумя путями: вегетативным и половым. Чтобы определить, какой из этих способов является более прогрессивным, нужно разобраться в их сути.
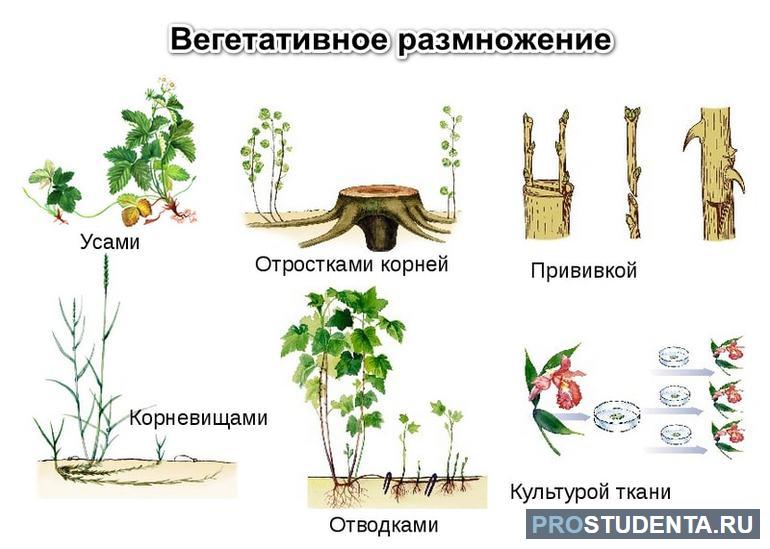
При вегетативном способе новое растение образуется из части тела родительской особи, например, из кусочков корня, луковиц, побегов, клубней. Это самый простой и быстрый путь размножения. Его часто используют при выращивании садовых культур, так как он дает большое количество урожая. Но вегетативный способ не позволяет получать различные комбинации генов у потомства. Растения, полученные таким путем, полностью наследуют все черты материнской особи.
Половой способ размножения предполагает слияние женских и мужских клеток, которые иначе называются гаметами. В результате этого процесса образуется зигота. Это клетка с двойным набором хромосом, из которой формируется зародыш. Он наследует признаки двух родителей. Молодое растение получает свойства, которые помогают ему выживать и адаптироваться к условиям внешней среды.
Части цветка
Характерной особенностью покрытосеменных растений является наличие цветка. Это орган размножения, который сформировался в ходе эволюции. Он представляет собой измененный побег, в котором происходит образование гамет и двойное оплодотворение у растений. В строении цветка можно выделить такие части:
- стерильные (околоцветник);
- репродуктивные (пестики, тычинки).
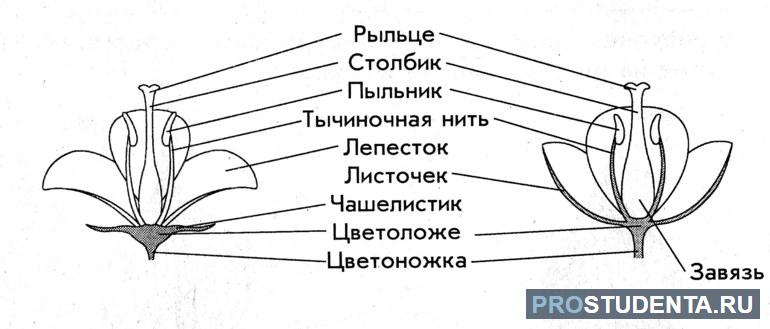
Околоцветник состоит из чашелистиков и лепестков, которые образуют чашечку и венчик. Стерильные части цветка не участвуют в размножении. Они предназначены для защиты репродуктивных органов — пестиков и тычинок.
Тычинка — это мужской орган размножения. В ее верхней части расположен пыльник. Он состоит из камер, в которых образуются мужские гаметы (пыльцевые зерна). Цветок может содержать от одной до сотен тычинок. Их количество зависит от разновидности растения.
Пестик — женский репродуктивный орган, расположенный в центре цветка. Здесь происходит процесс опыления и развитие зародыша. Пестичная часть цветка состоит из завязи, столбика и рыльца.
Мужские цветки содержат тычинки, а женские — пестики. Существуют также гермафродитные (обоеполые) растения. У них пестик и тычинки располагаются в одном цветке.
У однодомных растений тычиночные и пестичные цветки вырастают на одной особи. Это упрощает процесс размножения. Двудомные растения характеризуются тем, что мужские и женские цветки находятся на разных особях.
Образование мужских и женских клеток
Двойному оплодотворению предшествует формирование мужских и женских клеток. Этот процесс происходит в репродуктивных частях цветка.
В пыльниках тычинок расположены стволовые клетки. При делении они образуют микроспоры, из которых формируются пыльцевые зерна. Образование пыльцы происходит, когда лепестки цветка распускаются. Пыльцевое зерно состоит из двух видов клеток: генеративной и вегетативной. При делении генеративной клетки образуются две мужских гаметы, которые иначе называют спермиями. Они могут иметь разнообразную форму. Спермии не могут самостоятельно передвигаться, так как не снабжены жгутиками.
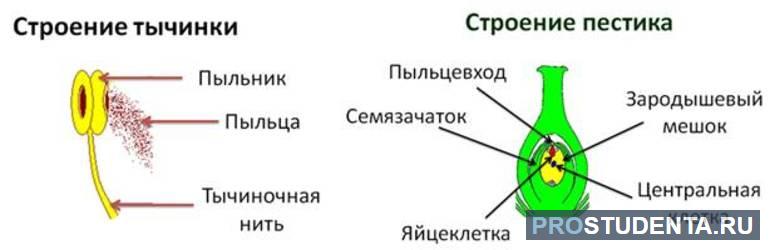
Внутри пестика формируются семязачатки. Они представляют собой выросты на стенках завязи. Каждая семяпочка содержит мегаспору. Это специальная клетка, которая в процессе деления дает начало яйцеклетке и зародышевому мешку. Так формируются женские гаметы в пестике.
Процесс опыления
Опылением называется перенос гамет с мужских цветков на женские. Это первый этап полового размножения. Пыльца переносится с тычинки на пестик разными способами. Биологи выделяют следующие разновидности этого процесса:

Самоопыление характерно для обоеполых растений. Перенос пыльцевых зерен происходит в пределах одного цветка. Пыльца падает на рыльце пестика под действием силы тяжести. В некоторых случаях ее перенос осуществляется ветром или мелкими насекомыми, обитающими внутри цветка. Обязательным условием самоопыления является одновременное созревание пестика и тычинок.
Двудомные и однодомные растения размножаются преимущественно перекрестным опылением. Это означает, что пыльцевые зерна переносятся с мужского цветка на женский. Этот процесс осуществляется с помощью ветра, воды или летающих насекомых. Важную роль в размножении растений играют пчелы. Одна особь способна осеменить за день около 2000−3000 женских цветков.
Разные виды цветковых растений имеют свои особенности. Это связано со способом переноса пыльцы. Растения, опыляемые насекомыми, отличаются крупными и ароматными цветками. Тычинки содержат крупные пыльцевые зерна, покрытые липким веществом. Это позволяет пыльце удерживаться на лапках насекомых.
Ветроопыляемые виды покрытосеменных выглядят более невзрачно. Их мелкая и сухая пыльца легко распространяется с дуновением ветра.
В некоторых случаях опыление производится вручную человеком. Это позволяет выводить новые сорта садовых и огородных культур, а также повышать урожайность. Чтобы искусственно оплодотворить большое количество растений, люди используют специальные устройства и инструменты.
Как происходит двойное оплодотворение
После опыления пыльцевые зерна попадают на рыльце пестика. Эта часть цветка выделяет липкую жидкость, которая помогает пыльце удерживаться.
Вегетативная клетка начинает делиться. Из неё образуется пыльцевая трубка, которая растет со скоростью 35 мм в час. Она прорастает через стенки рыльца и столбик пестика, а затем проникает в завязь, где находится семязачаток с яйцеклеткой и зародышевым мешком.
На конце пыльцевой трубки расположено два спермия. Мужские гаметы у растений не могут самостоятельно двигаться. Они перемещаются к семязачатку по мере роста трубки.
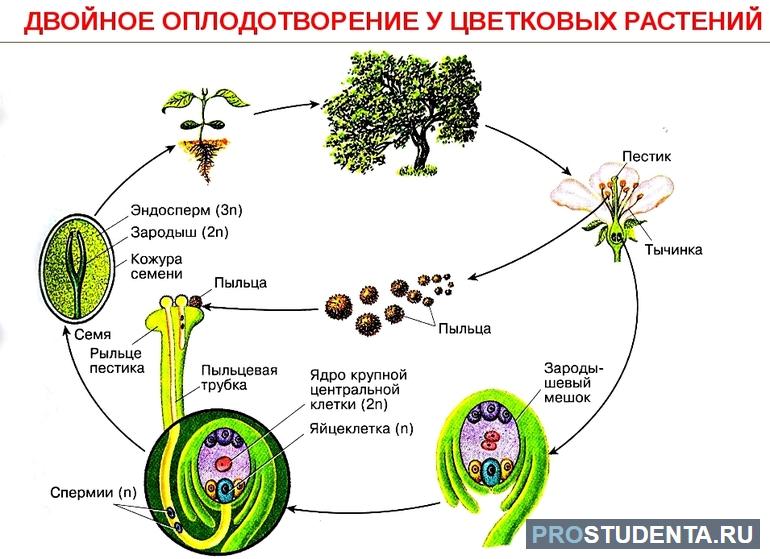
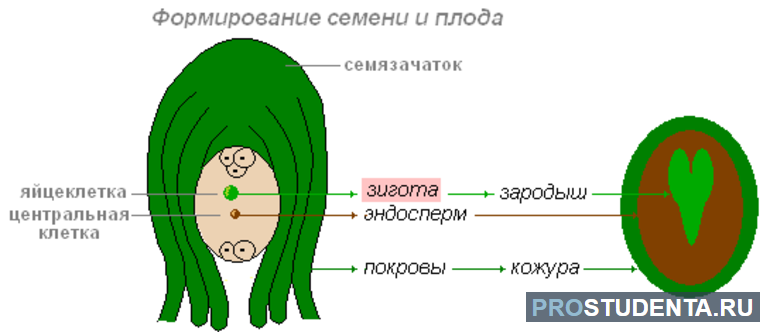
Семязачаток имеет специальное отверстие — пыльцевход. Через него прорастает пыльцевая трубка. Спермии попадают в зародышевый мешок семязачатка, где находятся яйцеклетка и центральная клетка. В этой полости происходит процесс двойного оплодотворения:
- Формирование зародыша. Один спермий оплодотворяет яйцеклетку. В результате образуется зигота, из которой формируется зародыш семени.
- Образование эндосперма. Второй спермий сливается с центральной клеткой зародышевого мешка. В результате оплодотворения формируется особый вид ткани — эндосперм.
Зигота и зародыш семени обладают двойным (диплоидным) набором хромосом, а эндосперм — тройным (триплоидным). Это связано с тем, что центральная клетка зародышевого мешка является двухъядерной. Эндосперм наследует хромосомный набор от двух ядер и спермия. В этой ткани накапливаются питательные вещества, которые необходимы зародышу. По мере развития семени завязь цветка превращается в плод.
Результатом опыления и оплодотворения является образование семян. С их помощью происходит размножение растений. Сложный процесс оплодотворения у покрытосеменных можно представить в виде схемы:
- формирование спермиев;
- перенос пыльцевых зерен с тычинок на пестик;
- прорастание пыльцевой трубки в семязачаток;
- оплодотворение яйцеклетки одним спермием;
- слияние второго спермия с центральной клеткой;
- развитие зародыша и эндосперма.
У голосеменных растений тоже образуются семена. Но у них не происходит двойного оплодотворения. Эндосперм голосеменных содержит одинарный набор хромосом. У этой группы растений не формируется плод, так как отсутствует цветок и его завязь.
Эволюционное значение
Биологи считают, что одновременное оплодотворение спермиями яйцеклетки и центральной клетки является прогрессивным способом размножения. Наличие эндосперма с тройным хромосомным набором создает наиболее благоприятные условия для формирования семян и плода.
Зигота и эндосперм развиваются одновременно. Зародыш получает питание на ранних этапах формирования. Семязачатку не приходится тратить ресурсы на создание запаса питательных веществ. Этот орган расходует свои силы исключительно на развитие зародыша.
Эндосперм у покрытосеменных разрастается быстрее, чем у голосеменных. Поэтому он дает развивающемуся зародышу больше питания.
У голосеменных растений эндосперм формируется даже тогда, когда зародыш отсутствует. У покрытосеменных ткань с запасом питания образуется только при развитии семени. Это позволяет экономить ресурсы и не тратить питательные вещества впустую.
Читайте также: