
Обратимые и необратимые повреждения растений реферат
Обновлено: 05.10.2024
Различают следующие основные виды повреждений клеток.
• Острое и хроническое.
• Парциальное, субтотальное, тотальное.
• Обратимое, необратимое.
• Специфическое, неспецифическое.
• Дисплазия.
• Дистрофия.
• Паранекроз.
• Некробиоз.
• Некроз.
Острое повреждение клеток под влиянием патогенных факторов и неблагоприятных внешних и внутренних условий возникает в течение секунд, минут, часов, дней, хроническое длится недели, месяцы, годы. При парциальном повреждении выходит из строя та или иная незначительная часть клетки, повреждения при этом обратимы, а клетка в целом остаётся жизнеспособной. При субтотальном повреждении страдает большая часть клетки с резким снижением её жизнеспособности. При тотальном наступает необратимая гибель всей клетки.
Обратимые повреждения после прекращения действия патогенного агента не приводят к гибели клеток. Возникающие при этом нарушения внутриклеточного гомеостаза обьгано незначительны и временны. Их можно устранить благодаря активизации внутри- и внеклеточных защитно-компенсаторно-приспособительных механизмов, что способствует восстановлению жизнедеятельности клетки.
Например, при воздействии эритемной дозы ультрафиолетовых лучей (вызывающей покраснение кожи) или кратковременном уменьшении кровотока (ишемии) в коже или во внутренних органах возникают незначительные, частичные изменения метаболизма, структуры и функций клеток, восстанавливающиеся через определённое время до нормального уровня.
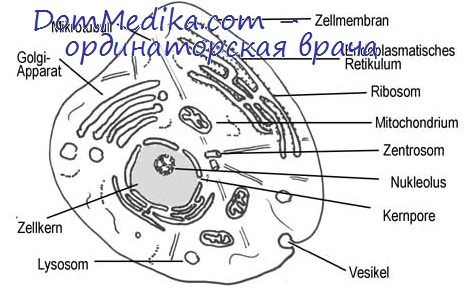
Необратимые повреждения клеток приводят к выраженным и стойким нарушениям внутриклеточного гомеостаза. Они не могут быть устранены даже максимальной активизацией защитно-компенсаторно-приспособительных механизмов в ещё оставшихся жизнеспособными внутри- и внеклеточных структурах.
Например, при длительных или резко выраженных влияниях на организм ультрафиолетовых и инфракрасных лучей, радиоактивных и токсических веществ, ишемии миокарда, головного и спинного мозга в различных клетках, тканях и органах организма возникают грубые расстройства структуры, метаболизма и функций, приводящие к снижению продолжительности их жизни и гибели (некрозу).
Гибель клеток обычно приводит к аутолизу за счёт выхода из лизосом и активизации гидролитических ферментов. Однако в ряде случаев гибель клеток и тканей может возникать без развития аутолиза. Это происходит, когда смерть целого организма наступает быстро (внезапно), например, при действии очень высоких или низких температур окружающей среды.
Специфические повреждения клеток характерны для того или иного конкретного патогенного фактора. К специфическим проявлениям повреждения клеток можно отнести:
• развитие иммунного гемолиза эритроцитов при наличии в организме антиэритроцитарных антител;
• угнетение активности холинэстеразы фосфорорганическими соединениями;
• подавление моноамин оксидазы ингибиторами и др.
Неспецифические (стереотипные) повреждения клеток возникают под влиянием различных патогенных факторов. К ним относят:
• увеличение объёма (набухание) клеток;
• угнетение энергетического и пластического обменов;
• нарушение метаболизма воды, электролитов, витаминов, липидов, углеводов, белков в результате развития ферментопатий;
• развитие ацидоза (первичного, вторичного);
• угнетение Na-, K-, Са-АТФазных насосов;
• увеличение внеклеточной концентрации калия и внутриклеточной концентрации кальция;
• нарушение структуры, метаболизма и функций клеточной и внутриклеточных мембран (прежде всего, повышение проницаемости, сорбционных свойств, изменение электропотенциала клеточных мембран, развитие ацидоза и др.).
Факторы окр. среды, действующие на растения, делятся на абиотические и биотические. По отношению к абиотич. факторам различ. три главных способа приспособления растений: механизмы, позволяющие избежать неблагопр. воздействие (переход в состояние покоя: образование почек, семян, вегетативных органов), специальные структурные приспособления (различные видоизменения листьев, стеблей цветков и т.д.), физиологические механизмы (С4-путь фотосинтеза, САМ-фотосинтез, увеличение вязкости цитоплазмы).
По отношению к биотич. факторам у растений также различ. несколько способов приспособл. (естественной защиты): приспособл. морфолог. характера, механизм неспецифического иммунитета, т.е. приспособл. биохимич. характера (фитонциды, фитоалексины, алкалоиды), механизм специфического иммунитет (выработка специализированных антител против возбудителей болезней).
Повреждение кл. может быть частичным или полным, обратимым или необратимым. Необр. поврежд. может привести к деструкции и гибели кл. Поврежд. кл. может быть первичным и вторичным. Первичное поврежд. кл. — это результат непосредственного действия повреждающего фактора. Различ. первичные повреждения: а) механические, б) термические, в) химические, г) радиационные. Вторичные поврежд. кл.— это такие, когда результат первичн. воздействия сам становится поврежд. фактором и вторично повреждает здоровые до этого момента структуры. Первич. поврежд. кл. факторы вызывают специфические, присущие только им эффекты. Эти эффекты связаны с характером первич. поврежд. фактора: а) механические — вызывают нарушение целостности структуры тк., кл., межкл. и субкл. структур. в) термические — связаны с денатурацией белков. белково-липидных комплексов и изменением вторичной структуры нуклеиновых кислот с) химические - угнетают активность ферментов, блокируют клеточные рецепторы, вызывают перестройку молекул за счет гидролиза, переаминирования и т.п. d) радиационные — приводят к разрушению молекул с образованием свободных радикалов. Ответная реакция поврежд. кл. стандартна и назыв. неспецифической реакцией клетки на повреждение. Причина такого стандартного ответа закл. в том, что при любом повреждении обязательно: 1) наруш. барьерные функции мембран клеточной и внутриклеточной; 2) выкл. ионные насосы. Основные структурные изменения следующие: а) повыш. прониц. мембраны пострадавшей кл.; б) уменьш. дисперсности коллоидов цитоплазмы и ядра в) увел. вязкости цитоплазмы г) увел. сродства цитоплазмы и ядра к ряду красителей.
По степени выраженности различ.: а) паранекроз — обратимые нарушения структуры и функции кл. б) некробиоз — необратимые поврежд. (гибель) части кл. в ткани. в) некроз — массовая гибель кл. с активацией лизосомальных ферментов и разрушением других кл. структур. Этот процесс называется аутолизом. Значение аутолиза — удаление мертвых клеток и замена их новыми клетками или элементами соединительной ткани.
ПРОЯВЛЕНИЯ поврежд. КЛ. 1. Ув. Прониц. цитоплазмат. мембраны: 1) белкам и коллоидным краскам (макромолекулы); 2) к аминокислотам и глюкозе (вещества с низкой молекулярной массой); 3) к ионам.
2. Умен. электрического сопротивления ткани.
Изменение физико-химических и функциональных свойств растительных клеток и тканей при повреждения и процессы адаптации; общие принципы адаптивных реакций растений на экологический стресс; стресс-белки.
Адаптация, т. е. приспособл. организма к конкретным усл. существования, у индивидуума достигается за счет физиологических механизмов (физиологическая адаптация).
Защита от неблагопр. факторов среды у растений обеспечив. особенностями анатомич. стр. (кутикула, корка, механические ткани и т. д), специальн. органами защ. (жгучие волоски, колючки), двигател. и физиологичес. реакциями, выработкой защ. вещ-в (смол, фитоалексинов, фитонцидов, токсинов, защитных белков).
Реакцию организма на любые отклонения от нормы называют стрессом (от англ. stress — напряжение). Самые разнообразные неблагопр. факторы могут действовать длительное время или оказывают сравнительно кратковременное, но сильное влияние. В первом случае, как правило, в большей степени проявляются специфические механизмы устойчивости, во втором — неспецифические.
У растений различают три фазы стрессовой реакции: 1) первичная стрессовая реакция, 2) адаптация, 3) истощения ресурсов надежности.

Факторы, способные вызвать стресс у растительных организмов, можно подразделить на три основные группы:
а) физические: недостаточная или избыточная влажность, освещенность или температура, радиоактивное излучение, механические воздействия;
б) химические: соли, газы, ксенобиотики (гербициды, инсектициды, фунгициды, промышленные отходы и др.);
в) биологические (поражение возбудителями болезней или вредителями, конкуренция с другими растениями, влияние животных, цветение, созревание плодов).
По отношению к засухе растения делятся на две группы: 1) пойкило-гидрические, не регулирующие свой водный режим и допускающие большую потерю воды (до воздушно-сухого состояния), не теряя жизнеспособности; 2) гомойогидрические растения, регулирующие водный обмен и отвечающие стрессом на водный дефицит.
Устойчивость растения к стрессовому воздействию зависит и от фазы онтогенеза. Наиболее устойчивы растения, находящиеся в покоящемся состоянии (в виде семян, луковиц и т. п.). Наиболее чувствительны — растения в молодом возрасте, в период появления всходов, в период формирования гамет.
К перв. Неспециф. процессам, происходящим в клетках растений при сильном и быстро нарастающем действии стрессора, отн. след.: пов. прониц. мембран, деполяризация мембранного потенциала плазмалеммы., вход Са 2+ в цитоплазму, сдвиг рН цитоплазмы в кислую сторону, активация сборки актиновых микрофиламентов и сетей цитоскелета, усиление поглощения О2, ускоренная трата АТР, развитие свободнорадикальных реакций,возрастание гидролитических процессов, активация и синтез стрессовых белков, увеличение синтеза этилена и АБК, торможение деления и роста, поглотительной активности клеток и других физиологических и метаболических процессо. Торможение функциональной активности кл. происх. в результате действия ингибиторов и переключения энергетических ресурсов на преодоление неблагопр. сдвигов.
Все эти явления адаптационного синдрома (стресса) взаимосвязаны и развиваются как каскадные процессы.
Особый интерес вызывают данные об активации в клетках в условиях стресса синтеза так называемых стрессовых белков с одновременным ослаблением синтеза белков, образ. в норм. усл. В ядре и в ядрышке белки теплового шока образ. гранулы, связывая матрицы хроматина, необх. для норм. метаболизма. После прекращения стрессового состояния эти матрицы вновь освобождаются и начинают функционировать. Кроме синтеза шоковых белков при неблагопр. обстоятельствах в кл. возрастает содер. углеводов, пролина, которые участвуют в защ. реакциях, стабилизируя цитоплазму. Благодаря своим гидрофильным группам пролин может образовывать агрегаты, которые ведут себя как гидрофильные коллоиды. Этим объясняется высокая растворимость пролина, а также способность его связываться с поверхностными гидрофильными остатками белков, что повыш. растворим. белков и защищает их от денатурации.
Механизмы стресса и адаптации на организменном уровне. На разных уровнях организации приспособление к экстремальным усл. осуществл. у растений неодинаково. Чем выше уровень биологич. организации (клетка, организм, популяция), тем большее число механизмов одновременно участвует в адаптации растений к стрессовым воздействиям.
На организменном уровне сохр. все механизмы адаптации, свойственные клетке, но дополняются новыми, отражающими взаимодействие органов в целом растении. Прежде всего это конкурентные отношения между органами за физиологически активные вещества и трофические факторы. Важнейший и очень характерный для растений механизм защиты от последствий действия экстремальных факторов — процесс замены поврежденных или утраченных органов путем регенерации и роста пазушных почек. Во всех этих процессах коррелятивного роста участвуют межклеточные системы регуляции (гормональная, трофическая и электрофизиологическая).
При неблагоприятных условиях существования в растениях резко возрастает выработка этилена и АБК, снижающих обмен веществ, тормозящих ростовые процессы, способствующих старению и опадению органов, переходу растительного организма в состояние покоя. Одновременно в тканях снижается содержание ауксина, цитокинина и гиббереллинов.
Замерзание
Зарождение льда, образование льда и его распространение
Температура плавления льда знают все: 0°C. На этой температуре даже основана шкала Цельсия.
А вот с температурой замерзания воды все сложно. Как бы странно это не звучало, при 0°C вода не всегда превращается в лед. Очень чистая, дистилированная вода может остаться жидкой вплоть до -40°C. Эту температуру называют точкой гомогенного зарождения льда. Вода, охлажденная до температур ниже 0°C, называется переохлажденной.
Дело в том, что для превращения в лед необходимы зародыши льда. Затем к зародышам присоединяются все новые молекулы воды, и вся вода превращается в лед. Проблема в том, что до -40°C зародыши льда быстро распадаются. Образование зародыша сопровождается выделением тепла, а выделенное тепло превращает кристалл обратно в воду.
Понимаю, что информация про -40°C может быть встречена в недоверием. Мы же видим лед на лужах или иней на траве даже после слабых заморозков. Ничего удивительного здесь нет, ведь в лужах вода не дистилированная. Зародыши льда легко образуются на взвешенных частицах, бактериях, поверхностях.
Есть еще одно важное свойства воды. Температура замерзания водных растворов уменьшается при увеличении концентрации соли.
В травянистых и древесных растениях лед обычно начинает образовываться при температурах от -2 до -4 °C.
В исследовании трех сортов черной смородины (Ribes nigrum) Картер и др. (1999) определили, что в веточках соцветий лед не образуется до -9°C, а в стебле лед образуется при температуре -2°C. Очевидно, что соцветия могут пережить заморозки за счет переохлаждения, и между стеблем и цветочной веточкой есть барьеры для распространения льда.
Процесс зарождения и распространения льда в растениях не до конца понятен. Методы исследования основаны на оценке количества зародышей льда в растении при разрушительном отборе образцов, или на использовании термопар. Эти методы, не могут определить ни первичные места зарождения, ни детали распространения льда по растению.
Недавно для исследования была использована инфракрасная видеотермография. Использование этой технологии для мониторинга замерзания основано на выделении тепла при переходе от жидкости ко льду.
Замораживание отделенного побега Picea pungens, наблюдаемое с помощью радиометра Inframetrics Model 760 Imaging Radiometer. A. Побег до замораживания. B. Замораживание побега начинается в нижней левой части при температуре около -5,9°C. C. Лед распространяется по побегу и попадает на хвою. D. Весь побег начал замерзать. Температура побега повышается примерно на 2°C в результате выделения теплоты плавления при образовании льда. Время от A до D 1 мин.
Замораживание отделенного побега Picea pungens, наблюдаемое с помощью радиометра Inframetrics Model 760 Imaging Radiometer. A. Побег до замораживания. B. Замораживание побега начинается в нижней левой части при температуре около -5,9°C. C. Лед распространяется по побегу и попадает на хвою. D. Весь побег начал замерзать. Температура побега повышается примерно на 2°C в результате выделения теплоты плавления при образовании льда. Время от A до D 1 мин.
Внеклеточное замерзание
При замораживании растения лед быстро распространяется во внеклеточном пространстве. Стенки клеток - эффективные барьеры для распространения льда. Поэтому, внутри клеток вода остается жидкой. Эта вода начинает выходить из клеток и присоединяться к растущим кристаллам льда. В результате, по мере снижения температуры клетки постепенно обезвоживаются, а внутриклеточный раствор становится все более концентрированным.
В этом плане действие мороза похоже на действие засухи.
Скорость роста кристаллов льда и обезвоживания клеток зависит от скорости охлаждения тканей. Когда скорость охлаждения медленная, клеточная вода постепенно переходит во внеклеточный лед. Концентрация внутриклеточного раствора постепенно увеличивается.
- температура замерзания внутриклеточного раствора уменьшается;
- раствору все сложнее покидать клетку из-за осмотического давления.
Понятно, что каждой температуре соответствует своя точка равновесия.
Однако, скорость охлаждения может оказаться слишком высокой, а скорость выхода воды из клеток - слишком низкой. В таком случае внутриклеточный раствор становится переохлажденным и склонным к образованию внутриклеточного льда. В природе на скорость охлаждения влияют изменение температуры, скорость ветра, воздействие солнечных лучей, масса тканей растения и характеристики теплопередачи. Быстрые колебания температуры бывают в холодные дни когда нагретая солнечным светом хвоя внезапно затеняется.
Переохлаждение
При быстром охлаждении растения внутриклеточная вода становится переохлажденной. Избегание замерзания путем переохлаждения - общий механизм выживания многих растений умеренной зоны.
Переохлажденная вода может превратиться в лед. Было замечено, что повреждение растений связано с замерзанием переохлажденной воды в клетках. Степень переохлаждения воды ограничена точкой гомогенного зарождения льда (-40°C), и, как ожидалось, выносливость тканей, в которых наблюдается переохлаждение, также ограничена.
Замораживание переохлажденной воды в растительных тканях может быть легко обнаружено с помощью дифференциального термического анализа (DTA). Ткани, в которых наблюдаются явления переохлаждения, обычно демонстрируют две температуры выделения тепла (экзотермы) при охлаждении. Первая экзотерма, или высокотемпературная экзотерма (HTE), возникает чуть ниже 0°C и связана с замерзанием внеклеточной воды. Вторая экзотерма, или низкотемпературная экзотерма (LTE), соответствует теплу, выделяемому при замерзании переохлажденной воды. LTE часто обнаруживаются вблизи -40°C, точки гомогенного зарождения льда и физического предела, до которого вода может быть переохлаждена до спонтанного образования льда. Проводя исследования лесной флоры в Скалистых горах Колорадо, Беквар и др. (1981) наблюдали LTE в веточках ели энгельмана (Picea engelmannii), пихты субальпийской (Abies lasiocarpa), сосне гибкой (Pinus fiexilis), можжевельнике обыкновенном (Juniperus communis), можжевельнике скальном (Juniperus scopulorum), псевдотсуге мензиса (Pseudotsuga menziesii) и пихте одноцветной (Abies concolor), но не обнаружили замерзания переохлажденной воды в ветках ели колючей (Picea pungens), сосны орегонской (Pinus ponderosa) и сосны скрученной (Pinus contorta). Беквар и др. (1981) также провели параллельные лабораторные испытания на замерзание и отметили, что повреждения от замерзания в древесных стеблях происходили в том же диапазоне температур, что и LTE у всех семи видов хвойных, которые демонстрировали переохлаждение.
Джордж (1982) сообщил, что небольшая часть воды (
Сакаи (1979) сообщил, что вода в вырезанных зимних почках нескольких хвойных видов переохлаждается до -17 - -35°C перед замерзанием. LTE были обнаружены в зимних почках Abies balsamea , Abies firma , Abies homolepis , Abies sachalinensis, Tsuga sieboldii, Tsuga diversifolia, Tsuga dumosa, Cedrus deodara, Keteleeria davidiana, Picea morrisonicola , Picea smithiana , Picea glehnii и Picea glauca. У каждого из этих видов LTE коррелировала с повреждением тканей при замораживании.
Повреждения при замерзании
Повреждение растений морозом - сложный процесс. На клеточном уровне на растение действует температура, механический стресс из-за льда в ткани, обезвоживание, вызванное замораживанием, и, как следствие, высокая концентрация внутренних растворителей. Однако принято считать, что при наличии внеклеточного льда основной причиной повреждения при замораживании является обезвоживание, вызванное замораживанием. Следовательно, повышенная устойчивость к стрессу обезвоживания была предложена в качестве основного результата закаливания у растений.
Необратимые и обратимые повреждения
Опыты с травами показали, что в зависимости от степени и продолжительности мороза полученное повреждение может быть либо необратимым (летальным), либо обратимым (ткани способны восстанавливаться после него и поэтому являются сублетальными).
Температура зарождения льда и скорость замораживания
При замораживании вода покидает клетки, внутриклеточный раствор становится более концентрированным, и это помогает ему оставаться жидким. Аналогичная картина возникает и при засухе. Соответственно, повреждения при замораживании зависят от способности клеток переносить обезвоживание, от температуры зарождения льда, скорости охлаждения и продолжительности воздействия холода.
Исследование с использованием парных срезов (две половинки) листочков картофеля (Solanum acaule) показало, что из двух половинок, замороженных до -3,5°C при скорости охлаждения 1°C/ч (медленное) или 3°C/ч (быстрое), последняя была необратимо и более серьезно повреждена, тогда как первая полностью восстановилась после мороза. Две половинки, замороженные до -1 или -3.5°C, продемонстрировали аналогичные повреждения, причем последняя была необратимо и более серьезно повреждена. Эти исследования показывают, что необратимые повреждения вызываются быстрым и сильным замораживанием тканей.
Почему половинки одного и того же листочка подверглись необратимому или обратимому повреждению, даже если они были заморожены до одинаковой температуры и, таким образом, испытали потенциально одинаковый уровень обезвоживания?
Вероятно, в случае необратимого повреждения тканей, клеткам не хватает времени, чтобы приспособиться к резким изменениям. Однако фактическое образование льда, а не просто температура ткани, определяет повреждение растительных тканей морозом. Например, в исследовании с листьями картофеля Линдстром и Картер (1985) не обнаружили никаких признаков повреждения после кратковременного охлаждения листьев даже до -6 °C, в то время как при наличии зародышей льда ткани сильно повреждались при -6°C.
Известно, что быстрое замораживание вызывает необратимые повреждения хвои, даже зимой, когда дерево очень устойчиво к морозу. Хвоя ели красной (Picea rubens), открытая солнцу с юга, получает более серьезные (необратимые) повреждения потому что быстрее охлаждается; хвоя ели красной на больших высотах на северо-востоке Северной Америки может охлаждаться со скоростью от 9 до 20°C в минуту из-за затенения и ветра. Снижение температуры на 10°C в минуту вызвало гибель хвои у туи западной (Thuja occidentalis) при температурах, которые в других условиях не вызывали повреждений. Перкинс и Адамс (1995) в эксперименте по контролируемому замораживанию отметили, что хвоя ели красной, замороженная со скоростью 2,6°C/мин не повредилась. Напротив, хвоя, замороженная при 11,9°C/мин (или >10°C мин), была необратимо повреждена и вызвала симптоматику, идентичную естественному зимнему повреждению на уровне ветвей, хвои и клеток. Иглы, замороженные при обеих скоростях охлаждения, демонстрировали HTE -4°C, но только быстро охлажденные иглы демонстрировали LTE -12°C, указывающую на внутриклеточное замораживание. Интересно, что у быстро замороженных игл (>10°C в минуту), но не подвергшиеся замораживанию ниже -10°C, не было видимых повреждений.
Повреждения морозом в природе
Хвойные могут повреждаться морозами даже в естественном ареале. Обычно это происходят в молодых, развивающихся и не закаленных тканях хвойных.
Повреждения весенними заморозками после появления молодых приростов часто встречаются по всей северной зоне распространения хвойных.
Повреждения также вызываются ночными заморозками летом, в период вегетации, когда активные и растущие ткани хвойных чувствительны даже к незначительному замерзанию.
Ночные заморозки могут повредить растения ранней осенью если погодные условия не способствовали закаливанию. Летом и ранней осенью, в основном, морозом повреждаются хвойные сеянцы и саженцы, а не крупные деревья.
Взрослые хвойные, растущие в суровом климате, обычно переносят самые сильные зимние морозы. Однако зимнее иссушение и снижение морозостойкости из-за теплых периодов могут вызвать повреждения тканей хвойных зимой и ранней весной.
Заморозки вызывают два процесса.
Радиационные заморозки случаются в спокойные ясные ночи когда поверхность земли охлаждается за счет излучения тепла в атмосферу.
Адвекционные заморозки случаются когда воздух, охладившийся ниже точки замерзания в другом месте, поступает на участок.
Масштаб адвекционных заморозков бывает разным: от крупномасштабной фронтальной активности (холодная арктическая воздушная масса) до локальных нисходящих потоков плотного холодного воздуха и небольших скоплений холодного воздуха на пологой местности.
Нильсен и Роулунд (1996) сообщили о повреждениях в недавно созданных и молодых насаждениях ели сихтинской (Picea sitchensis) в Дании. Развивающиеся побеги были нормальными до начала июня, когда они внезапно завяли, а затем погибли. Заморозки повредили камбий в конце апреля, когда температура внезапно понизилась после теплого периода в марте.
Похожие повреждения произошли в насаждениях ели сихтинской в Шотландии (Gregory and Redfern 1987). Эффективная сумма температур (сумма среднесуточных часовых температур выше +5°C) в конце мая была вдвое выше средней эффективной суммы температур за предыдущий 30-летний период, из-за чего почки распустились рано и повредились заморозком.
Летние и ранние осенние заморозки являются обычным явлением в высоких широтах и на больших высотах. Растения чувствительны даже к небольшим (-3°C) заморозкам в летний период. Повреждения летними заморозками могут ограничиться временным угнетением фотосинтеза, но сильный свет после заморозков может еще больше повредить фотосинтетические системы и клеточные органеллы. Обесцвечивание хвои после летних заморозков сильнее проявляется у сеянцев и саженцев, находящихся под прямыми солнечными лучами, чем у затененных. Это было продемонстрировано на саженцах сосны обыкновенной и ели обыкновенной в Швеции, а также на саженцах ели черной и лиственницы американской (Larix laricina), растущих на торфяниках в центральной Альберте.
Многие ели были повреждены в южной и центральной Швеции зимой 1984-85 годов. Повреждения наблюдались как в еловых насаждениях, так и в средневозрастных древостоях. Зима 1984-85 гг. характеризовалась холодной и дождливой осенью и умеренными температурами в ноябре-декабре, за которыми последовало быстрое снижение температуры в январе. Таким образом, недостаточно закаленные деревья могли начать вегетацию в ноябре и декабре, когда температура была выше нормы, а повреждения возникли из-за быстрого снижения температуры в январе.
Необычайно суровая зима случилась в Скандинавии зимой 1986-87 гг. После этой зимы наблюдалась преждевременная потеря хвои у сосны обыкновенной в юго-восточной части финской Лапландии. Сильные морозы в декабре 1986 и январе 1987 года привели к повреждению корней, поскольку температура почвы была исключительно низкой из-за неглубокого или отсутствующего снежного покрова. В центральной части Финляндии зимой 1986-87 гг. также наблюдались повреждения морозом. Среднемесячная температура в январе была примерно на 12°C холоднее чем рассчитанное среднее значение за 30 лет.
Зимнее иссушение - это распространенная форма повреждения морозом хвойных деревьев зимой и ранней весной. Зимнее иссушение или повреждение морозной засухой происходит когда надземные или расположенные выше линии снега части деревьев подвергаются воздействию теплого, сухого воздуха и солнца, в то время как земля и часть стебля остаются замороженными. Иссушение случается не только на замерзших, но и на незамерзших холодных почвах.
Чем холоднее почва и чем дольше период промерзания почвы, тем больше у растений повреждений от иссушения.
На иссушение влияет и скорость испарения воды через поверхность хвои. Даже низкая скорость потери воды может привести к сильному, накапливающемуся за зиму водному стрессу. Таким образом, толщина защитного слоя хвои (кутикулы) - важным фактором в предотвращении зимнего иссушения. На толщину и устойчивость кутикулы влияют летние температуры и зимние ветры.
От зимнего иссушения могут страдать даже деревья в своем естественном ареале обитания.
В настоящей статье подробно изложена суть процесса повреждения клетки. Описаны два возможных варианта рассмотрения данного процесса: в качестве причины и следствия развития патологического процесса. Представлены основные классификации повреждений: по степени нарушения внутриклеточного гомеостаза, по происхождению и другие. Разобраны возможные причины клеточных повреждений. Также сделан акцент на возможности рассмотрения повреждения с точки зрения вреда и пользы для организма.

2. Патологическая анатомия: учебник / А.И. Струков, В.В. Серов. – 5-е изд., стер. – М.: Литтера, 2010. – 880 с.: ил.
3. Патофизиология: учебник : в 2 т. / под ред. В.В. Новицкого, Е.Д. Гольдберга, О.И. Уразовой. - 4-е изд., перераб. и доп. - ГЭОТАР-Медиа, 2017 - Т. 1 - 848 с.: ил.
Введение.
Повреждение клетки - изменения структуры, метаболизма, свойств и функций, которые ведут к нарушению жизнедеятельности [1]. Повреждения можно рассмотреть с двух сторон: как причину и следствие развития патологического процесса в организме. К примеру, если наблюдаются повреждения гепатоцитов печени, то возможно развитие патологических процессов в органе с последующим нарушением его нормальных функций. При возникновении инфаркта миокарда будет нарушаться нормальная деятельность клеток с их последующей гибелью в результате возникновения гипоксии сердечной мышцы; будут происходить нарушения со стороны кровоснабжения организма, что станет причиной нарушения функционирования других клеток и возникновения осложнений на системы органов. В любом случае, задача организма состоит в ликвидации клеточного повреждения. Достигнуть этого можно путём включения всевозможных защитных, компенсаторных клеточных механизмов, которые будут стараться уменьшить неблагоприятное действие повреждающего фактора (по возможности остановить его действие) и устранить все последствия его воздействия на клетку. Однако для того, чтобы ликвидировать повреждение, необходимо разобраться какие виды повреждений бывают, какие факторы их вызывают, в какой форме они проявляются и какое воздействие оказывают на состояние организма в целом.
Результаты.
Классификация клеточных повреждений.
По степени нарушения внутриклеточного гомеостаза выделяют [2]:
- Обратимое повреждение. Оно возможно, когда различные защитные механизмы способны устранить последствия повреждения. К примеру, у эритроцитов при кратковременном изменении осмотического давления крови.
- Необратимое повреждение. Повреждение будет необратимым, если привлечения всевозможных защитных и компенсаторных механизмов клетки недостаточно для ликвидации последствий повреждения. Это происходит, когда повреждающий стимул оказывает влияние на клетки слишком долго и/или слишком интенсивно. Это наблюдается при длительной ишемии сердечной мышцы, в процессе которой происходит повреждение кардиомиоцитов необратимого характера. В целом можно сказать, что обратимые повреждения будут нарушать гомеостаз организма, ухудшать его общее состояние и нормальное функционирование, а необратимые - приводить к гибели клеток и сильно снижать продолжительность их жизни.
Патогенетические виды повреждений [3]:
1.Повреждение насильственного характера — наблюдается в случае, когда клетка абсолютно здорова, ее защитно-компенсаторные механизмы находятся в норме, однако сила и интенсивность повреждающего агента настолько велика, что он, воздействуя на клетку, приводит к её повреждению. Наиболее чувствительны к этому повреждению функционально малоактивные клетки, которые обладают небольшой силой собственных механизмов для поддержания гомеостаза.
2.Повреждение цитопатического характера — наблюдается в случае, когда клетка слабая и её механизмы защитно-компенсаторного характера нарушены. В такой ситуации даже воздействие повреждающего фактора обычной силы приведёт к повреждению клетки. К цитопатическому характеру нарушения будут относится те виды повреждений, которые возникают из-за отсутствия недостающих и нужных клетке компонентов. (Например, гипоксическое повреждение - при недостатке кислорода). Клетки, чья реактивность и функциональная активность высокая в обычных для них ситуациях (например, нейроны) будут в большей степени чувствительны к этому виду повреждения.
Причины клеточных повреждений по происхождению [4]:
Внешние(экзогенные) стимулы воздействуют извне. Выделают следующие виды агентов экзогенного происхождения:
1)Агенты физического характера: колебания температур, действие электрического тока, механические повреждения, ультрафиолет, электромагнитные явления.
2)Агенты химического характера: кислоты/щелочи, соли тяжелых металлов, низкомолекулярные неорганические вещества (альдегиды, фенолы), высокомолекулярные вещества (иммуноглобулины).
3) Агенты биологического характера: вирусы, бактерии.
Внутренние(эндогенные) стимулы находятся в клетке. Выделают следующие агенты эндогенного происхождения:
1) Агенты физического характера: свободные радикалы, смена осмотического давления
2) Агенты химического характера: ионы, кислород, метаболиты, физиологически активные вещества (гормоны)
3) Агенты биологического характера: продукты жизнедеятельности других поврежденных и погибших клеток
Клеточное повреждение под действием одного повреждающего агента, может быть связано с увеличением или уменьшением его количества в клетке. К примеру, рассмотрим избыток и недостаток кислорода в тканях: при его избытке активируется процесс свободнорадикального перекисного окисления липидов (СПОЛ), в результате которого повреждаются ферменты и клеточные мембраны; недостаток кислорода, наоборот, нарушает нормальное течение окислительных процессов, снижает образование АТФ и, в целом, наблюдается нарушение со стороны функций клеток.
Выводы.
В заключение хотелось бы отметить, что повреждение клеток вне зависимости от вида, всегда будет иметь как отрицательные проявления, так и положительные. Если вспомнить суть первичного и вторичного повреждения клетки, то нельзя однозначно сказать, хорошо это или плохо, так как для разных ситуаций повреждение клетки будет по-разному сказываться на общее состояние организма. К примеру, после первичного повреждения клетки, организм не смог своевременно его ликвидировать, и оно перешло во вторичное повреждение, распространившись на соседние клетки [5]. С одной стороны, это плохо, так как на заживление вторичного повреждения организму придется затратить больше сил, энергии, времени и включить все свои дополнительные защитные и компенсаторные резервы. Но, с другой стороны, если соседние клетки оказываются опухолевыми, то в результате перехода первичного повреждения во вторичное, эти клетки будут также повреждаться и удаляться, соответственно, это будет положительным эффектом.
Читайте также: