
Появление семян у семенных папоротников
Обновлено: 05.10.2024
Изучая палеозойские и раннемезозойские растения, находимые в земных слоях в виде отпечатков и окаменелостей, палеоботаники нередко встречают ископаемые листья, по строению неотличимые от перистых листьев настоящих папоротников, но несущие семена на своих верхушках или по бокам жилок.
Такое удивительное сочетание признаков семенных растений и папоротников, внервые установленное в 1904 г. выдающимися английскими палеоботаниками Ф. Оливером и Д. Скоттом, и дало повод назвать эту группу растений семенными папоротниками (Pteridospermae).
Это была обширная группа растений, из которой к настоящему времени описано несколько сот видов, объединяемых в четыре основных порядка — лигиноптерисовые (Lyginopteridales), медуллозовые (Medullosales), кейтониевые (Caytoniales) и глоссоптерисовые (Glossopteridales) и около 10 семейств.
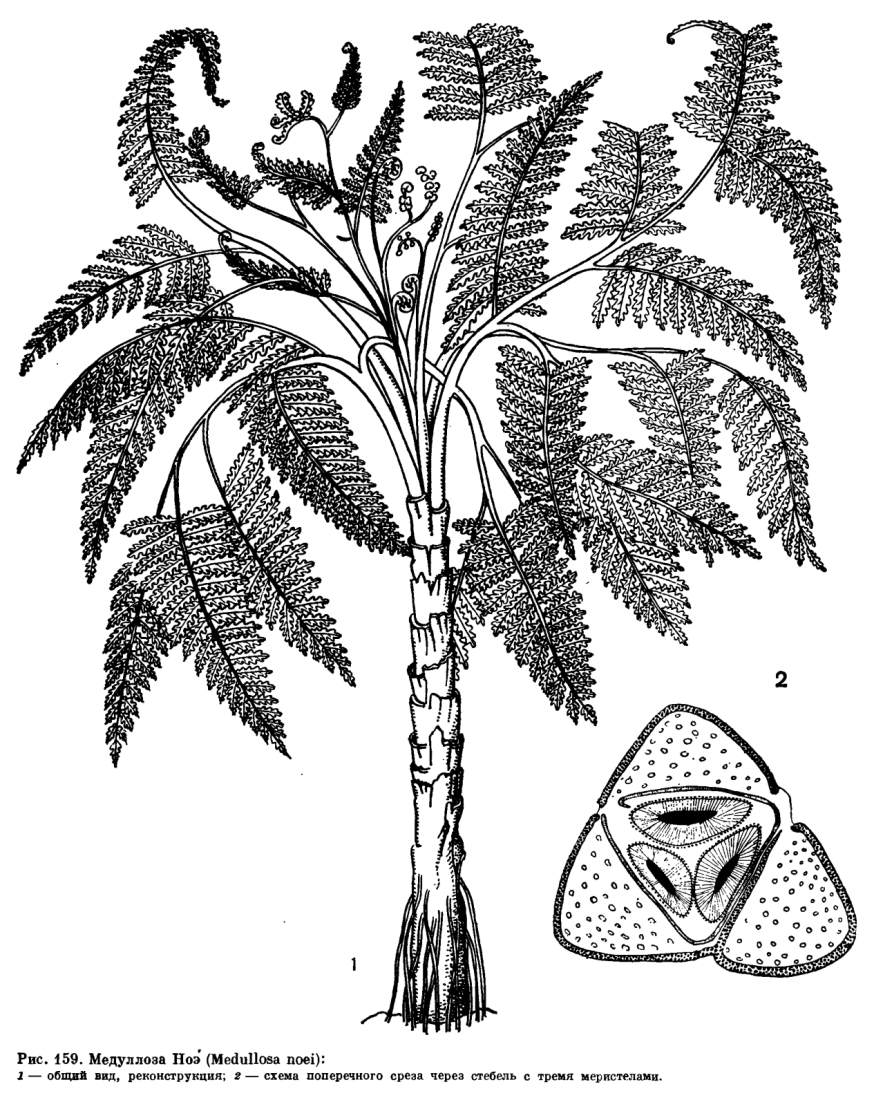
Внешний вид этих вымерших растений наиболее удачно передает реконструкция медуллозы Ноэ (Medullosa noei), выполненная американскими палеоботаниками В. Стюартом и Т. Делевориасом по отпечаткам листьев и срезам окаменелых стеблевых частей (рис. 159).
Семенные папоротники — полностью вымершие растения. Они существовали с позднего девона до раннего мела. Наиболее часто их остатки встречаются в отложениях карбона и перми. Одни из них были распространены по всему земному шару, другие, как, например, ангароптеридиум (Angaropteridium), были свойственны только ангарским флорам, а глоссоптерисовые и користоспермовые, напротив, приурочены главным образом к районам, соответствующим древнему материку Гондване, который существовал с середины карбона до конца триаса.
Наиболее распространено мнение, что семенные папоротники карбона были обитателями более возвышенных участков, обрамлявших приморские заболоченные леса. Однако все больше данных за то, что, например, заключительные этапы существования формации карбопового низинного заболоченного леса, как правило, сопровождались пышным развитием семенных папоротников или кордаитовых.
Семенные папоротники были похожи на настоящие папоротники не только по строению перистых листьев, но и по внешнему облику растений: древовидных, лиановидных и травянистых.
Стебли древовидных семенных папоротников были или прямыми, или дихо- и моноподиально ветвящимися. У одних листья сидели в очередном порядке на радиально-симметричном стебле, у других были сближены на верхушке и располагались в виде кроны, как у многих современных древовидных папоротников. Поверхность стебля многих растений была покрыта волосками, а иногда особыми железками, как, например, железками булавовидной формы у калиматотеки (Calymmatotheca), которые выполняли, по-видимому, секреторную функцию. У одних растений стволы были более или менее гладкие, с рубцами от опавших листьев; у других покрыты основаниями опавших листьев, как у медуллозы. У лигиноптерисовых (Lyginopteridaceae) стебли были диаметром 2—4 см, а у медуллозовых (Medullosaceae) их диаметр достигал 20 см. У некоторых семенных папоротников на стебле между листьями и в узлах располагались воздушные корни. Стебли пельтаспермовых (Peltaspermaceae) и кейтониевых (Caytoniaceae) до сих пор не найдены, и обе группы известны только по находкам стерильных и фертильных листьев и семян.
Листорасположение у семенных папоротников было спиральным. Как у лигиноптерисовых, так и у медуллозовых листья были довольно крупными и в основании вильчато делились на две симметричные половины, каждая из которых была перисто рассечена. В почкосложении листья были улиткообразно закручены, как у современных и многих ископаемых папоротников.
У растений из порядка глоссоптерисовых листья сидели спирально или мутовчато (глоссоптерис) в утолщенных узлах тонких стеблей. Они были довольно крупными, но очень различными по форме — цельные, цельнокрайние, линейные, ланцетные, лопатчатые, яйцевидные, сидячие и на коротких черешках. Если у более древнего рода гангамоптерис (Gangamopteris) не было ясно выраженной средней жилки, то у глоссоптериса (Glossopteris) она была развита хорошо.
Размножение семенных папоротников происходило с помощью семян, в которых до сих пор не найдены зародыши. Отсутствие зародыша, констатированное при изучении многих сотен ископаемых семян этих растений, до сих пор представляется загадкой. Существует точка зрения, согласно которой семенные папоротники не имели настоящего семени, хотя у них был семязачаток. В связи с этим их, а также современные саговниковые и гинкговые причисляли не к семенным, а к так называемым семязачатковым растениям. Однако едва ли это решает вопрос. Несомненно, что зародыш должен был развиться до прорастания этого семязачатка. По-видимому, развитие зародыша у семенных папоротников происходило в семязачатках, уже опавших с материнского растения на почву, как это происходит нередко у ныне живущего голосеменного гинкго двулопастного.
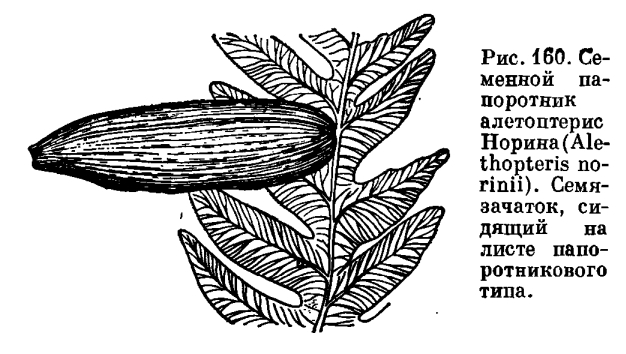
Семязачатки развивались на расчлененных папоротниковидных листьях, располагаясь или по краю, или на верхушке листа (рис. 160). У наиболее примитивных родов семязачатки сидели на концах боковых или верхушечных перышек (сегментов), у более подвинутых родов (например, у сфеноптериса тонкого — Sphenopteris tenuis) — на поверхности перышек. Иногда положение семязачатков было вторичноверхушечным за счет редукции и видоизменения перышек в результате их превращения в семяножку.
У большинства семенных папоротников семязачатки были окружены так называемой купулой, т. е. особым покровом, имеющим у различных растений очень разное строение и форму, иногда чашевидную, конусовидную или дисковидную. Считается, что купула возникла из сегментов (долек) листа, окружающих семязачаток. Листовое происхождение купулы подтверждается ее строением у наиболее древних представителей группы.
Семязачатки были одиночными, как у лигиноптерисовых, или многочисленными, как у глоссоптерисовых. У лигиноптерисовых они достигали в длину 5,5 мм, у пельтаспермовых— 7 мм, а у користоспермовых — 3,5—7 мм.
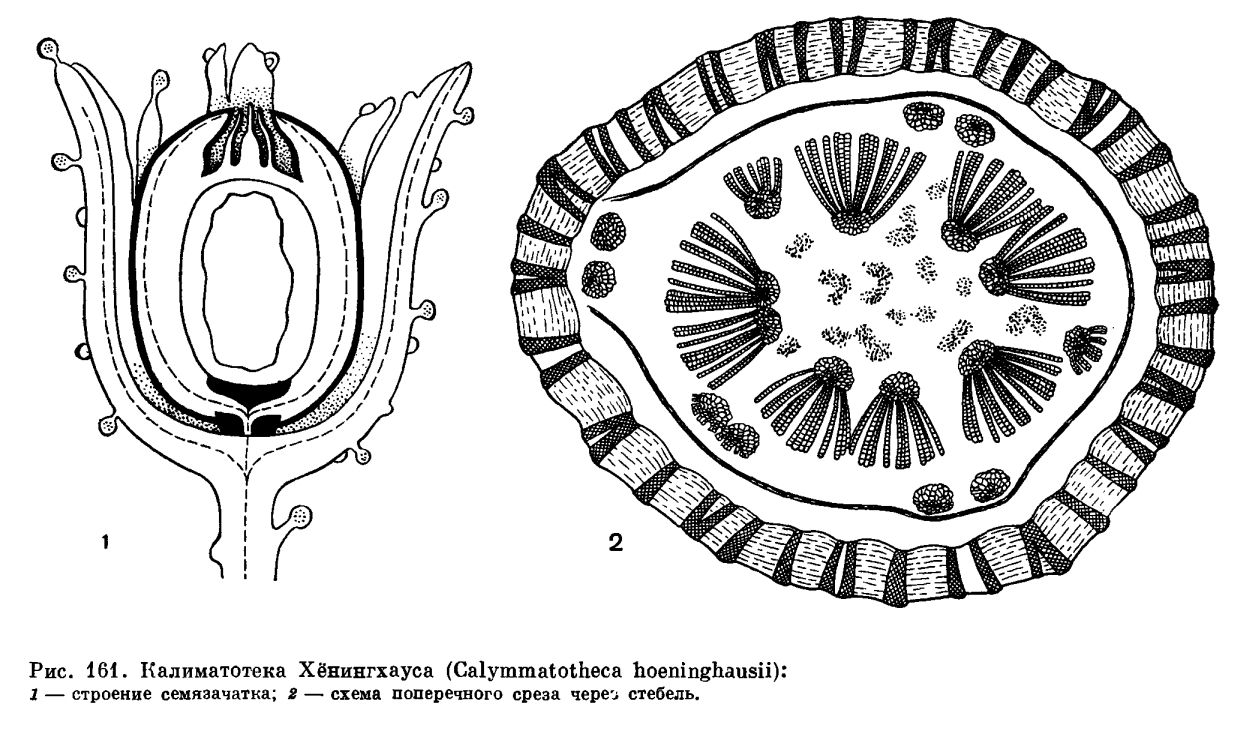
Рассмотрим строение семязачатка на примере калиматотеки Хёнингхауса (Calymmatotheca hoeninghausii, рис. 161). Снаружи семязачаток окружен купулой, несущей железки. Из ножки купулы проводящий пучок переходит в ножку семязачатка, имеющего овальную форму. По строению проводящий пучок ножки сходен с пучком листового черешка. Наружный покров семязачатка (интегумент) возникает, по мнению ученых, в результате длительной эволюции из кольцеобразно расположенных спорангиев, которые, потеряв фертильность и срастаясь между собой, обеспечили защиту центральному, сохранившему фертильность мегаспорангию. Таким образом, семязачаток — это синангии. Подтверждение этому — большое количество примитивно устроенных семязачатков, у которых сохранились признаки синангиальной структуры. Нуцеллус, соответствующий мегаспорангию, обычно тесно спаян с интегументом, но иногда, оставаясь свободным в верхушечной области семязачатка, он образует так называемую колонку пыльцевой камеры, в которой нередко находят пыльцу, причем иногда сохранившуюся в проросшем состоянии. На этом основании сделано предположение, что пыльцевая камера была заполнена сахаристой жидкостью, которая являлась благоприятной средой для прорастания пыльцы. Центральная часть семязачатка занята большой функционирующей мегаспорой, от которой у семенных папоротпиков чаще сохраняется только оболочка, но нередко также ткань гаметофита с архегониями, в которых даже сохраняются яйцеклетки.
Мужские спороношения у растений этой группы — обычного папоротникового типа. Они представлены или краевыми микроспорангиями, как у лигиноптерисовых, или микросинангиями, иногда сложно устроенными, как у медуллозовых и кейтониевых. Пожалуй, наиболее интересны сложно устроенные и очень разнообразные по форме мужские спороношения медуллозовых.
Микроспоры семенных папоротников были разными по величине и строению. Например, у пельтаспермовых они были очень мелкими, диаметром всего 30 мкм, однобороздными, а у лигиноптерисовых и медуллозовых — диаметром до 400—500 мкм, и имели одну дистальную борозду наряду с трехлучевым тетрадным рубцом. Проросшие микроспоры этого типа были найдены в пыльцевых камерах некоторых семян, в частности, у пахитесты (Pachytesta). Предполагают, что гигантские микроспоры переносились на семязачатки, видимо, еще до полного развития мужского гаметофита, который завершал свое развитие в питательной жидкости пыльцевой камеры семязачатка.
Семенные папоротники — группа наиболее примитивная среди голосеменных растений. Одни ученые приходят к выводу, что они занимают промежуточное положение между настоящими папоротниками и голосеменными, в то время как другие считают эти группы возникшими и эволюционировавшими параллельно. Как уже сказано, листья семенных папоротников часто очень похожи на листья настоящих папоротников или даже неотличимы от них. Это представляет яркий пример параллельного развития гомологичных органов. Но наличие семян, анатомическое строение стебля и корня и способность семенных папоротников к вторичному росту достаточно резко отличают их от настоящих папоротников.
Анатомическое строение стеблей и корней семенных папоротников очень специфично. Так, стебель молодого растения калиматотеки был протостелическим, но с возрастом он становился сифоностелическим с хорошо развитой сердцевиной и гнездами склеренхимных клеток (рис. 161). Вокруг сердцевины располагались многочисленные мезархные пучки, их метаксилема состояла из крупных (в сечении) лестничных, сетчатых и точечных трахеид. Последние несли многорядные окаймленные поры на всех стенках. Как у большинства семенных папоротников, у калиматотеки была довольно мощная вторичная ксилема, состоящая также из точечных трахеид, но с порами только на радиальных стенках. Довольно широкие древесинные лучи, состоящие из паренхимных клеток, разбивали вторичную ксилему на секторы. Интересной особенностью внутреннего строения стебля этого семенного папоротника является структура наружной коры, которая состояла из сложного сетчатого переплетения тяжей склеренхимных клеток. На тангентальном срезе они имели вид ячеистой стенки, а на поперечном — вид радиально вытянутых полос, имеющих форму трабекул. Петли сетки были заполнены паренхимной тканью. Листовые следы имели двухпучковое строение. В основании листа эти два пучка срастались в F-образный (на поперечном срезе) пучок.
У медуллозовых строение как стебля, так и листовых черешков было принципиально иным. Их стела рассечена на ряд элементарных стел (меристел), каждая из которых развивала свои вторичные проводящие ткани — ксилему и флоэму (рис. 159). Число меристел у видов медуллозы было различным. Хотя меристелы имеют вид самостоятельных проводящих систем, они анастомозируют друг с другом, соединяются в узлах, затем снова расходятся. Наружная кора у медуллозовых была обычно более просто устроена, чем у лигиноптерисовых, и состояла из массивной паренхимной ткани, в которой были рассеяны одиночные секреторные каналы, как у мараттиевых и саговниковых. Меристелы были погружены в основную ткань стебля, представленную паренхимой, в наружной части которой проходили многочисленные листовые следы. Листовые следы в своей нижней части близ стелы стебля имели концентрическое строение, а в наружной зоне коры и в листовом черешке были представлены многочисленными коллатеральными пучками. Этим они очень напоминают стебли однодольных растений.
Остатки семенных папоротников играют большую роль в сложении каменных углей СССР, Западной Европы и Северной Америки. Почти на каждом срезе через угольные почки, составляющие окаменевшие участки древнего торфяника, превращенного в уголь, можно найти стебли, корни, семена и пыльцу семенных папоротников.
Группа семенных папоротников взята на вооружение биостратиграфами, занимающимися вопросами расчленения палеозойских и мезозойских отложений. Например, по появлению отпечатков листьев растения каллиптерис (Callipteris) проводится нижняя граница пермского периода.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .

Задание 16 № 25296
Установите соответствие между примерами и путями эволюции: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
А) появление семян у семенных папоротников
Б) редукция пищеварительной системы у ленточных червей
В) формирование ловчего аппарата у венериной мухоловки
Г) формирование роющих конечностей у медведки
Д) возникновение теплокровности у млекопитающих
3) общая дегенерация
Запишите в таблицу выбранные цифры под соответствующими буквами.
1) ароморфоз: А) появление семян у семенных папоротников; Д) возникновение теплокровности у млекопитающих;
2) идиоадаптация: В) формирование ловчего аппарата у венериной мухоловки; Г) формирование роющих конечностей у медведки;
3) общая дегенерация: Б) редукция пищеварительной системы у ленточных червей; Е) отсутствие листьев у растения повилики.
Ароморфоз — прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации организмов.
Идиоадаптация, одно из главных направлений эволюции, при котором возникают частные изменения строения и функций органов при сохранении в целом уровня организации предковых форм.
Семенные папоротники – древняя вымершая группа Голосеменных, обнаруженная в палеозойских и раннемезозойских отложениях, их возраст – около 350 млн лет (рис.3, 4).

Рис. 3. Семенной папоротник – медуллоза
1 – реконструкция внешнего облика; 2 – поперечный разрез черешка.

Рис. 4. Семенной папоротник – калимматотека
А – общий вид (часть растения); Б – репродуктивный побег (микроспорофиллы); В – семязачаток (снаружи покрыт плюской); Г – продольный разрез через семязачаток и плюску: 1 – вегетативная часть побега, 2 – репродуктивная часть, 3 – спорангии.
По внешнему облику эти растения были близки к папоротникам, но имели семязачатки, которые располагались прямо на листьях, что и дало повод назвать эту группу Семенными папоротниками. После оплодотворения семязачаток отделялся от растения. Далее, без покоя уже на земле развивался спорофит. Поэтому их нередко называют семязачатковыми, а не семенными. Наиболее изучены рода Medullosa и Calymmatotheca.
Класс Саговниковые
Включает один порядок Саговники и одно семейство Саговниковые, объединяющие около 130 видов тропических и субтропических вечнозеленых невысоких деревьев, иногда кустовидных или эпифитных форм Юго-Восточной Азии, Средней Америки.
Современные виды саговников — это остатки когда-то пышной саговниковой флоры мезозойской яры. Появились они в пермском периоде палеозоя.
Внешне напоминают древовидные папоротники или пальмы. Для них характерно наличие стержневого корня, типичной эустелы стебля, камбиального кольца со вторичными камбиальными элементами коры. Однако, как и у папоротников, у саговников имеется сильно развития крупноклеточная сердцевина, богатая крахмалом. Листья саговников крупные, перистораздельные, жесткие.
Один из типичных представителей саговников — саговник поникающий (Cycas revoluta). Это двудомное растение с коротким стволом, покрытым панцирем из бурых жестких остатков оснований листьев.
Семена саговников крупные (от 3—4 до 8 см в длину), с сочным красным или оранжевого цвета покровом. Семена съедобны для многих видов животных (обезьян, медведей, диких свиней, грызунов). Среди классов Голосеменных саговники имеют самые примитивные по строению семена. Это выражается в формировании у них мощной дифференцированной семенной кожуры, обильного эндосперма, недоразвитости зародыша в морфологически и физиологически зрелом семени.
Из середины ствола, коры и эндосперма семян добывают крахмал — саго. Саговники декоративны и ценятся в озеленении.
Класс Беннеттитовые
Беннеттитовые вымерли около 70 млн лет назад в период расселения Цветковых. А появились в начале мезозоя (около 200 млн лет назад). По своему облику они напоминали Саговниковые, и длительное время их ископаемые остатки относили к Саговниковым. Однако они значительно отличаются: наряду с однополыми, как у Саговниковых, стробилами (р. Williamsonia), у Беннеттитов имелись и обоеполые (р. Williamsoniella, Cycadeoidea). Это единственная группа, в которой есть представители со стробилами, объединяющими микро- и макроспорофиллы.
В стробиле Cycadeoidea на мясистом семяложе располагались многочисленные семязачатки, разделенные стерильными межсеменными чешуями. К основанию семяложа прикреплялись перистые микроспорофиллы со многими микроспорангиями; снаружи стробилы были защищены многочисленными густо опушенными покроволистиками (рис.5).

Рис. 5. Беннеттитовые
А – реконструкция внешнего облика: 1 – вильямсония; 2 – цикадеоидея; 3 – вильямсониелла; Б – репродуктивные органы: 1 – разрез через стробил цикадеоидеи; 2 – разрез через женскую часть стробила; мсп – микроспорофилл; п – периант; спф – мегаспорангиофор; сз – семязачаток; ст.ч – стерильная чешуя; 3 – продольный разрез через семя; з – зародыш.
Такой стробил отдаленно напоминал цветок Покрытосеменных, в котором макроспорофиллы (пестики) и микроспорофиллы (тычинки) расположены примерно там же. Поэтому многие ботаники рассматривали Беннеттиты как возможных предков Цветковых растений.
Класс Гинкговые
Гинкго двулопастное – это эндемик горных лесов Китая (рис. 6). В настоящее время культивируется в садово-парковых посадках Восточной Азии, Северной Америки и Европы (может произрастать до широты Киева). Это высокое (до 30 м) листопадное дерево с характерными вееровидными двулопастными листьями с дихотомическим ветвлением. Растение двудомное. Микростробилу (мужскую шишку) образуют микроспорофиллы, сидящие спирально на длинной оси (микростробила) в виде тонкой ножки, на конце которой висят 2 (3-4) микроспорангия. Макростробилы состоят из длинной ножки, заканчивающейся двумя семязачатками, из которых обычно только один развивается в семя с мясистым покровом.
Семязачаток гинкго по своему строению напоминает семязачаток саговника и ненамного отличается от него (рис. 7).

Рис. 6. Гинкго двулопастной
А – побег мужского растения с сережковидной шишкой; Б – микроспорофилл с микроспорангиями; В, Г – побеги женского растения с шишками; Д – женская шишка (разрез); Е – продольный разрез верхней части семязачатка; Ж – семя, З – продольный разрез семени:
1 – интегумент (сочная оболочка); 2 – нуцеллус; 3 – археспориальная камера; 4 – архегоний; 5 – эндосперм; 6 – наружная мясистая часть оболочки; 7 – каменистая часть; 8 – тонкий пленчатый слой оболочки; 9 – зародыш.
Развитие гаустории, процесс оплодотворения, формирование зародыша и семени в целом также похожи на саговниковые. Если опыление происходит весной, то оплодотворение осуществляется только осенью, иногда уже в опавших семязачатках, ни внешне, ни по размерам не отличающихся от созревших семян. Они имеют внешний мясистый слой семенной кожуры, срединный каменистый (склеротеста) и внутренний, похожий на пергамент. Семена прорастают без видимого периода покоя, что относится к числу примитивных признаков.
Класс Хвойные
Наиболее многочисленная группа среди современных Голосеменных, геологическая история которой восходит к раннему карбону.
Хвойные – включают около 560 видов, 56 родов и 7 семейств. Основное видовое разнообразие приходится на южное полушарие: умеренные области Новой Зеландии, Австралии, Южной Америки – бассейн Тихого океана. Чем это объясняется? В этой части планеты не было резких колебаний климата, как в континентальных районах.
Стебель имеет тонкую кору и массивную древесину, которая на 90-95 % состоит из трахеид и очень мало паренхимы. У многих Хвойных и в коре, и в сердцевине имеются смоляные каналы, наполненные эфирными маслами. У большинства Хвойных развивается мощный стержневой корень, от которого отходят длинные боковые корни. Листья большинства Хвойных игловидные, узколинейные или чешуевидные и называются хвоей. Однако у тропического рода Agathis (сем. Araucariaceae) они напоминают листья Однодольных – широкие ланцетные (длина 18 см, ширина 6 см), также у представителей семейства Подокарповые (длина 35 см, ширина 9 см).
Вечнозеленые, плотные, более или менее жесткие, кожистые листья Хвойных содержат крупные смоляные каналы. Листья жесткие с толстой кутикулой, устьица погружены в углубления, которые заполнены воском, что уменьшает испарение.
Микростробилы, часто называемые мужскими колосками, состоят из укороченной оси, на которой расположены сильно редуцированные микроспорофиллы. На каждом микроспорофилле образуется от 2 микроспорангиев (у сосновых) до 7 (и даже 15 – у агатиса и араукарии).
У большинства Хвойных мужские колоски мелкие (1-3 см), но у араукарии они достигают 25 см в длину и 4-5 см в диаметре, напоминая стробилы Саговников.
Семена сильно различаются по величине и форме. У некоторых родов они снабжены одним большим крыловидным придатком, у других – 2-3 небольшими крыльями.


Семенные папоротники или птеридоспермы существовали с карбона по пермь включительно и поначалу отпечатки их листьев принимались за обычные или истинные папоротники, размножающиеся спорами. Со временем, по мере увеличения количества находок, они были выделены в отдельную группу ископаемых растений. Наиболее известные получили родовое имя медуллозовые (Medullosales). Их древовидные стволы имели необычную форму в виде параллельных трубок, каждая из которых имела проводящую ткань, обеспечивающую влагой большой сложный лист, напоминающий лист папоротника. Эти растения были небольшой высоты, где-то около 3 метров, занимали средний ярус в лесном биоценозе и размножались семенами.

Medullosa — ствол, характерный для таких известных семенных папоротников, как Neuropteris, Alethopteris,Odontopteris

Схема, показывающая устройство листовых пластин некоторых из семенных пепоротников
Далее представлены находки с отпечатками семенных папоротников, найденные на породных отвалах Донецка

Два рахиса Neuropteris шахта им. Горького, Донецк

Пиннулы (листовые пластины) Neuropteris под микроскопом

Пиннулы (листовые пластины) семенных папоротников под микроскопом

Листовая пластина Neurodontopteris (размер листа 10х7 мм), отвал шахты Центрально-Заводская

Macroneuropteris — отвал шахты им. Шверника

Neuropteris ovata, окрестности г. Константиновка

Alethopteris grandini, отвал шахты №12 наклонная, Донецк

Macroneuropteris, окрестности г. Константиновка

Типичная смесь листьев птеридоспермов из лесной подстилки

Neurodontopteris, отвал шахты Центрально-Заводская

Reticulopteris munsteri, отвал шахты Центрально-Заводская

Листья Neuropteris

Отпечатки семян птеридоспермов — Trigonocarpus

Trigonocarpus

Trigonocarpus

Лист Odontopteris (Mixoneura), отвал шахты Моспинская

Фруктификация семенного папоротника, отвал шахты Моспинская

Alethopteris lonchitica, отвал шахты им. Шверника, Донецк

Mariopteris muricata, отвал шахты №7-8, Донецк

Mariopteris muricata, отвал шахты Моспинская

Отпечаток листьев Mariopteris, отвал шахты Моспинская

Папоротники относятся к наиболее древним споровым растениям. Обитают в самых разнообразных экологических условиях: в заболоченных местах и водоемах, лесах тропического и умеренного климата. Наиболее известные представители - щитовник мужской, орляк, страусник. В жизненном цикле папоротников, хвощей, плаунов доминирует спорофит.
Расцвет папоротников произошел около 358 млн лет назад и длился примерно 65 млн лет, в честь папоротников назван период палеозойской эры - карбон, или каменноугольный период, который длился указанный промежуток времени. Именно папоротникам принадлежит активная роль в углеобразовании: в карбоне леса заселяли древовидные папоротники, достигавшие 40 и более метров в высоту.

Древесные формы папоротников сохранились до наших дней, однако большинство представителей - травянистые растения, у которых отсутствует камбий, а значит, нет и вторичной древесины.
Папоротники относятся к группе сосудистых растений, так как имеют жилки - сосудисто-волокнистые пучки, в отличие от мхов, которые не имеют жилок и не являются сосудистыми растениями. Механическая прочность обеспечивается отложением склеренхимы вокруг проводящих пучков (жилок).

В отличие от мхов, папоротники имеют проводящую ткань в стеблях и корнях, состоящую из ксилемы и флоэмы. Заметьте, я написал "корень" - у мхов не было и корней, вместо них были ризоиды, выполняющие аналогичную корням функцию. Корни папоротников, хвощей и плаунов всегда придаточные и растут от видоизмененного побега - корневища.
Строение
Рассмотрим строение папоротников на примере типичного представителя - щитовника мужского. Это широко распространенный папоротник, типичный для климата умеренных широт. Представляет собой многолетнее травянистое корневищное растение.
Образован пучком сильно расчлененных листьев, отходящих от корневища. Нарастают листья верхушкой, образуя завитки - "улитки".

Отметьте, что лист папоротника называется вайя (от греч. baion - пальмовая ветвь). В отличие от настоящих листьев, вайя имеет недетерминированный верхушечный рост. Листья имеют черешок, прикрепляющийся к стеблю, который может продолжаться в рахис - главную ось сложного листа, соответствующую центральной жилке.

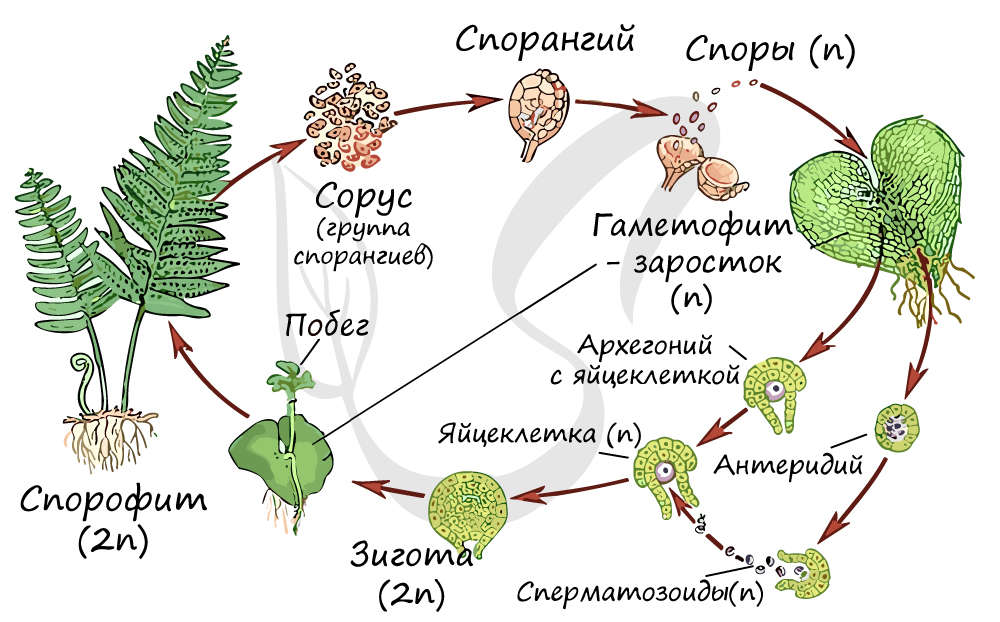
Жизненный цикл папоротниковидных
Листостебельное растение папоротника, изображенное выше - спорофит (2n). Спорофит доминирует в жизненном цикле папоротниковидных, в отличие от цикла мха, где спорофит, по сути, является придатком гаметофита (редуцирован). На нижней стороне вайи располагаются спорангии, собирающиеся в сорусы - группы близкорасположенных спорангиев. На спорофите (2n) в спорангии после мейоза образуются споры (n).

В 2012 году группа ученых под руководством Ксавьера Нобли из Университета Ниццы выяснили, что спорангий имеет специальный механизм "катапульты", споры из него вылетают со скоростью около 10 м/с.
Гаплоидные споры (n) прорастают в заросток (n), небольшую пластинку (несколько мм) сердцевидной формы. Заросток зеленого цвета, способен фотосинтезировать и прикрепляется к почве ризоидами. На нем образуются мужские и женские половые органы - соответственно антеридии и архегонии. Сперматозоид (n), образовавшийся в антеридии, благодаря воде (во время дождя) попадает в архегоний, где сливается с яйцеклеткой (n) и образуется зигота (2n).
Из зиготы развивается зародыш, который проникает в ткани архегонии с помощью особого приспособления - гаустории (от лат. haustor - черпающий, пьющий). Гаустория представляет собой ножку, внедряющуюся в ткани заростка и поглощающую из него питательные вещества. Начинается бурный рост зародыша, образуется побег, а затем взрослое растение - спорофит (2n). Цикл замыкается.
Значение папоротников
Папоротники являются главным компонентом многих лесных сообществ, звеном в цепи питания - продуцентами (производителями органических веществ). Человек использует папоротник в декоративных целях. Молодые побеги некоторых папоротников съедобны и употребляются в пищу: побеги орляка, листья страусника обыкновенного.
Щитовник мужской имеет медицинское значение: из его корневищ изготавливают противогельминтный препарат.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: