
При прорастании семян необходимым фактором для индукции синтеза хлорофилла является
Обновлено: 05.10.2024
Фитогормоны - это вещества, вырабатывающиеся в процессе обмена веществ и оказывающие в ничтожных количествах влияние на физиологические процессы.
Ауксины - это вещества индольной природы. Основным фитогормоном типа ауксина является β-индолилуксусная кислота (ИУК) она синтезируется из аминокислоты триптофана. Образование идет в меристематических тканях. Передвигаются из верхушки побега вниз к его основанию, а далее от основания корня к его окончанию. Передвижение идет по проводящим пучкам. Транспорт ИУК происходит как пассивно, так и активно.После окончания реакции гормон должен быть разрушен или инактивирован (фермент ИУК-оксидаза). Кроме того, ИУК может переходить в неактивное состояние образуя комплексы с сахарами, аминокислотами и другими веще-ствами. Такие комплексы могут распадаться, и тогда активности гормона быстро восстанавливается. С помощью комплексов в органе образуется запас гормона.
1)ИУК стимулирует растяжение клеток. Механизм заключается в активации протонных пом, закислении апопласта, снижении прочности клеточной стенки, поступлении воды и растягивании. Вызванное ауксином увеличение поступления воды приводит к быстрому растяжению клетки, уменьшению вязкости цитоплазмы и изменению скорости ее движения, что влияет на скорость химических реакций.
2) стимулирует синтез ДНК, РНК, белка.
3) стимулирует образование придаточных корней .
4) влияет на энергетический обмен.
5) стимулирует прорастание семян.
6)Под влиянием ауксина активируются кальциевые каналы что способствует транспорту воды и питательных веществ к месту действия гормона. ИУК: клетки и ткани, обогащенные ауксином, становятся как бы центрами притяжения этих веществ. Это приводит к усиленному росту органов.
Таким образом, ауксины определяют направление транспорта веществ в растении. Регулируя распределение и транспорт веществ, ауксины определяют полярность растения. 7)Асимметричное распределение ИУК вызывает неравномерный рост стеблей, корней, результатом которого являются движения органов растения.
Гиббереллины - это большая группа соединений, чтобы различать их, им присвоили определенные номера: ГК1, ГК2, ГК3 и т. д. Наиболее распространен гиббереллин ГК3 - гибберелловая кислота С19Н22О6 . По химической природе гиббереллины - это тетрациклические дитерпеноиды, состоящие из четырех остатков изопрена. Синтез происходит путем превращения мевалоновой кислоты . Место синтеза - молодые листья, а в клетке - пластиды. Транспорт в растении не является полярным. Гиббереллины свободно передвигаются из листьев вверх и вниз. по ситовидным трубкам, паренхимным клеткам, по сосудам транспортируются в связанной форме - гиббереллин-глюкозиды. Инактивация гиббереллинов происходит в результате их превращения в гиббереллин-глюкозиды и сложные эфиры. Разрушаются с помощью ферментов. Мех-м действя. Наиболее типичное действие гиббереллинов - это стимуляция роста стебля Рост стеблей вызван только растяжением клеток. Гиббереллин действует и на эмбриональную фазу роста клетки, а также стимулирует рост столонов и задерживает образование клубней. Гиббереллины накапливаются в семенах, луковицах, почках, клубнях. Они активизируют гидролитические ферменты и их синтез в зерновках злаков, что вызывает более быстрые превращения запасных веществ и прорастание семян. Гиббереллины – стимулируют цветение ряда растений.
Цитокинины - производные аденина, у которого аминогруппа в шестом положении замещена различными радикалами. Синтезируются из мевалоновой кислоты. Распространяется цитокинин вверх с ксилемным током (пассивно и неполярно). Синтез цитокининов происходит в разных участках растения в зависимости от этапа онтогенеза. Концентрации этих гормонов происходит благодаря соединению цитокининов с глюкозой. Мех-м действия. Главные функции цитокининов стимуляция клеточного деления (усиливают синтез белка и нуклеиновых кислот, активируют рост клеток) и дифференцировки, а также задержка процессов старения.
Обладают огромной аттрагирующей способностью, т. е. притягивают ассимиляты (аминокислоты, углеводы) и регуляторные вещества к клеткам и тканям, содержащим большое количество этого гормона. Задерживают старение листьев вызывая образование новых тилакоидов гран и стромы в хлоропластах, разрушившихся при старении, усиливает синтез хлорофилла. Нарушают покой, например свежеубранных клубней картофеля, активируют прорастание семян, активируют открывание устьиц, В культуре ткани стимулируют деления клеток. Подавляют рост боковых корней. Снимают апикальное доминирование, вызванное ауксинами. Способствуют росту бессемянных плодов.
К ингибиторам относят абсцизовую кислоту (АБК), этилен, брассиностероиды и др.
АБК содержится в корнях, стеблях, почках, листьях, плодах, во флоэмном и ксилемном соке, но особенно много ее в покоящихся почках, семенах, клубнях. Синтезируется из мевалоновой кислоты, Транспортируется АБК по сосудам и ситовидным трубкам вверх и низ во все органы. Физиологическая роль. АБК тормозит все процессы роста: задер-живает растяжение и деление клеток у молодых проростков и в культуре ткани; ингибирует распускание почек. Вызывает покой у семян. АБК играет роль антитранспиранта(закрывает устица). АБК также способствует запасанию гидратной воды в клетке.
Этилен. Он хорошо растворим в воде, поэтому может транспортироваться в водном растворе. Образуется в плодах, цветках, листьях, облиственных стеблях, корнях и семенах в наибольшем количестве этилен образуется в стареющих или созревающих тканях. Синтезируется из метионина. Этилен легко окисляется.
Физиологическая роль. Этилен в норме абсолютно необходим для созревания плодов, синтезируясь в клетках самого плода. Для начала созревания его концентрация должна увеличиться в три раза. Этилен - гормон механического стресса. Этилен стимулирует увядание цветков, опадение листьев и плодов, так как он ингибирует синтез и передвижение ауксина, подавляющего опадение и вызывает образование отделительного слоя в основании черешка или цветоножки, т.е. этилен регулирует явление листопада. Способствует заживлению ран (под действием этилена активизируется особая ткань - раневая перидерма.). Тормозит рост главного и стимулирует образование боковых корней.
Брассиностероиды образуются в незрелых семенах. Подавляет образование придаточных корней. Повышают устойчивость растений к низким и высоким температурам, дефициту влаги в почве и воздухе.
Применение фитогормонов в практике растениеводства.
Ауксины:1. Для усиления корнеобразования у черенков, восстановления корневой системы при пересадке растений. 2. Для образования партенокарпических плодов, повышения урожая томатов и некоторых других культур. 3. Для предохранения плодов от предуборочного опадения. 4. Для ускорения прорастания семян некоторых растений. 5. Применяются как селективные гербициды.
Гиббереллины.1. Для усиления роста стебля конопли, сахарного тростника. (увеличивается выход волокна конопли с гектара). 2. Для повышает урожай зеленой массы кормовых бобов.
Этилен-продуценты применяют: для ускорения созревания плодов, ускорения опадения листьев - дефолиации. Для индукции цветения. Для борьбы с полеганием зерновых культур в посевах.
Синтез хлорофилла
Биосинтез хлорофилла, все ферменты которого локализованы в хлоропластах, подразделяется на следующие этапы:
δ-Аминолевулиновая кислота → Порфобилиноген → Уропорфириноген I и III копропорфиноген III → протопорфиноген IХ → протопорфирин IХ → Mg-протопорфирин IХ метиловый эфир Mg-протопорфирина → протохлорофиллид → протохлорофиллид голохром → хлорофиллид а голохром → хлорофиллид а → хлорофилл.
1 стадия – исходным веществом в биосинтезе порфиринов является сукцинил Со-А (из лимоннокислого цикла) и глицин, из них образуется δ-аминолевулиновая кислота. Этот путь характерен для микроорганизмов и животных. Реакция идет с участием фермента – АЛК-синтаза:
Существует мнение, что АЛК в хлоропластах образуется другим путем, вероятнее всего из интактной молекулы глутамата. Фермент, катализирующий реакцию, удалось выделить из пластид в чистом виде недавно; реакция представляет собой новый внутримолекулярный перенос аминогруппы:
В последнее время уточнили, что δ-аминолевулиновая кислота образуется из С 5-дикарбоновых кислот: глутаминовая кислота через 2-гидрооксиглутаровую преобразуется в 4,5-диоксивалериановую, которая затем амминируется за счет аланина и других кислот.
Показательно, что это выраженное разветвление путей биосинтеза гема у животных и высших растений происходит, вероятно, на ключевой стадии биосинтетического пути – образовании АЛК.
Интересно было бы выявить каким из путей синтезируются молекулы АЛК, дающие начало тем небольшим количествам производных гема, которые выявлены у растений, выращенных в темноте: имеет ли место рассмотренный путь синтеза из глутамата или здесь действует путь, который происходит у животных (но его трудно выявить, поскольку в реакциях участвуют очень малые количества субстрата)?
2 стадия – образование первичного пиррола: 2 молекулы δ-АЛК → порфобилиноген превращаются в присутствии фермента АЛК-дегидратазы. Порфобилиноген – это первый предшественник металлопорфиринов, имеющий пиррольную природу.
3 стадия – образование циклического тетрапиррола. Четыре молекулы порфобилиногена превращаются в уропорфиноген I, а затем III:


4 стадия происходит отщепление 4 СО 2от остатков СН 2СООН → СН 3 в положениях 1, 3, 5, 8 молекулы уропорфириногена с участием фермента уропорфириногендекарбоксилазы и образуется копропорфириноген:

5 стадия – копропорфиноген превращается в протопорфириноген IХ за счет окислительного декарбоксилирования в 2 и 4 положениях:

6 стадия –в результате реакции дегидрирования, при которой отщепляются 6Н + из протопорфириногена IХ образуется протопорфирин:
7 стадия – происходит включение Mg (NН → N) и образуется Mg-протопорфирин IХ.
Вероятно, на стадии образования протопорфирина IХ происходит также разветвление путей биосинтеза – один ведет к образованию железопорфиринов (гем), другой – к магнийпорфиринов (хлорофиллам). В растениях обнаружены оба пути, у животных – только первый.
В биосинтезе железосодержащих порфиринов участвует фермент феррохелатаза, который выделен как из пластид, так и митохондрий растений. Этот фермент эффективно преобразует протопорфирин в протогем (гем b), который входит в состав цитохромов, каталазы, пероксидазы и гемоглобина. Нужно отметить, что химически встроить Mg 2+ в протопорфирины намного труднее, чем Fe 2+ . Какой фермент катализирует включение Mg 2+ в молекулу протопорфирина пока неизвестно.


8 стадия – превращение Mg-протопорфирина IХ в монометиловый эфир Mg-протопорфирина. Происходит этерификация метиловым спиртом. Фермент-S-аденозил-L-метионин-Mg-протопорфирин-метилтрансфераза осуществляет перенос СН 3в 6 положение от S-аденозил-L-метионина. Далее, окисление пропионата в положении с образованием группы С=О.
9 стадия – образование протохлорофиллида происходит в результате этирификации (этирификация – образование сложных эфиров из кислот и спиртов) карбоксильной группы метанолом, замыканием пятого (фуранового) кольца окончательным формированием боковых цепей: в 4 положении винильная группа СН=СН 2восстанавливается до этильной группы С 2Н 5, получается Mg-винилфеопорфирин а 5-протохлорофилл (ид).
Как и хлорофилл протохлорофиллид находится в связанной с белком форме.
10 стадия – в результате неферментативной индуцируемой светом реакции восстановления в кольце IV (появляются два атома Н) протохлорофиллид превращается в хлорофиллид (это хлорофиллы, в молекуле которых нет терпеноидной – обычно фитольной – боковой цепи). Спектр действия хлорофиллида подобен спектру поглощения протохлорофиллида, источником водорода в этой реакции служит белок (возможно НАДФ – донор водорода). Выделен в чистом виде фермент НАДФН – протохлорофиллид-оксиредуктаза. Таким образом, восстановление до хлорофиллида катализируется светом и происходит в пигмент-белковом комплексе, который имеет название протохлорофиллид-галохрома.
11 стадия – последняя стадия синтеза хлорофилла – образование сложного эфира хлорофиллида с фитолом, которая происходит в липидной фазе хлоропластов, поскольку фитол не растворяется в воде, с участием фермента хлорофиллаза. Фитол – полиизопреновое соединение синтезируется с ацетил-СоА через мевалоновую кислоту.
Хлорофилл bотличается, как мы отмечали от хлорофилла атолько тем, что боковым заместителем при С 3является группа СНО вместо СН 3. Соотношение хлорофиллов а/ bв пределах одного вида – величина довольно постоянная.
Несмотря на большие усилия исследователей, пока никому не удалось детально выяснить, каким путем происходит биосинтез хлорофилла b(его окисление).
Синтез хлорофилла зависит от генетических факторов. В результате генных мутаций появляются растения альбиносы, которые живут пока имеют запасные вещества из семян. Пятнистость – результат отсутствия синтеза хлорофилла в некоторых частях листа, что также зависит от генов.
Содержание хлорофилла в листе колеблется очень мало. Это связано с тем, что старые молекулы пигмента беспрерывно разрушаются и образуются новые. При этом один процесс уравновешивает другой.
Кроме света, синтез хлорофилла зависит и от условий минерального питания. Прежде всего, необходимо железо, которое выполняет каталитические функции. При недостатке железа листья желтеют. Большое значение имеет обеспеченность растений азотом и магнием, так как эти элементы входят в состав молекулы пигмента. При нехватке меди хлорофилл легко разрушается.
Этиолированные проростки содержат небольшое количество протохлорофиллид – белкового комплекса (голохром), и при кратковременном освещении в них происходит быстрое стехиометрическое восстановление протохлорофиллида до хлорофиллида, который затем медленно этирифицируется и преобразуется в хлорофилл а. Если проростки затем вновь возвратить в темноту, в них образуется приблизительно такое же количество протохлорофиллида, которое было вначале, и при осветлении он тоже превращается в хлорофиллид. Таким образом, при освещении этиолированных проростков короткими вспышками света (по 10 –4 с), которые чередуются с 10–15 минутными темновыми промежутками, можно накопить большие количества хлорофилла.
Хотя у большинства высших растений синтез хлорофилла происходит только на свету, некоторые из них, например, сеянцы хвойных растений, могут синтезировать хлорофилл в темноте. Способны к темновому синтезу хлорофилла и многие водоросли, например бурые и сине-зеленые. Пока что непонятно, почему последняя стадия восстановления пигмента у этих организмов не зависит от освещения (света). Однако показано, что развивающиеся семядоли хвойных синтезируют значительное количество хлорофилла в темноте только до той поры, пока они находятся в контакте с макрогометофитом, т. е. до того времени пока макрогометофит не исчезнет. Семядоли, которые отделяют от макрогометофита, даже если их поместить в питательный раствор, синтезирует значительно меньше хлорофилла. Следовательно, какое-то вещество из макрогометофита непосредственно участвует в темновом синтезе хлорофилла.
Реакцией, на уровне которой происходит начальный этап биосинтеза хлорофилла, является синтез аминолевулиновой кислоты (рис. 2.6).
АЛК – это субстрат первого основного (решающего) этапа в синтезе тетрапиррола. Это значит, что, вероятно, АЛК лимитирует скорость всего процесса. Об этом свидетельствует и ряд данных, например, добавление АЛК к этиолированным проросткам в темноте приводит к увеличению количества протохлорофиллида (ПХД) по меньшей мере в 10 раз.
Рис. 2.6.Регуляция биосинтеза порфиринов в развивающихся хлоропластах. Стрелки () показывают чем ингибируется процесс
Считают, что протохлорофиллид или другой контролируемый посредник подавляет синтез фермента (или ферментов), действия на стабильную mРНК.
Поскольку при биосинтезе хлорофилла и биосинтезе гема используется один и тот же накопленный промежуточный продукт, можно думать, что регуляция происходит именно в той точке, где пути биосинтеза расходятся, т. е. на том этапе, на котором атом металла встраивается в молекулу. Вероятно, протохлорофиллид регулирует включение Mg, а так же, как отмечали, синтез белков.
Синтез каротиноидовначинается с ацетил-CоА через мевалоновую кислоту, геранилгеранилпирофосфат до ликопина. Ликопин (С 40Н 56) – является предшественником всех других каротиноидов. Синтез идет в темноте, но ускоряется при действии света. Ликопин имеет ациклическую структуру (отсутствие кольца с двух концов молекулы) и характерен для плодов томата. Циклизация в концах (одного или двух) молекулы ликопина приводит к образованию разных колец (β- и ε-ионовых). Так образование двух β-ионовых колец (по обоим концам молекулы) приводит к образованию β-каротина. Вообще, последовательность превращения каротиноидов следующая:
β-каротин (два β-иононовых кольца) → α-каротин (одно β-, другое ε-иононовое) → лютеин (ε-иононовые кольца и ОН-группа в кольце при С 3);

β-каротин → зеаксантин (2 кольца β-иононовые и группа ОН в кольцах) → виолаксантин (два β-иононовых кольца, кроме ОН-групп появляются эпоксигруппыв каждом кольце).
Фотопротекторная роль каротиноидов, т. е. защита хлорофилла от фотоокисления, еще раз подчеркнем, объясняется их способностью взаимодействовать с возбужденными молекулами кислорода и хлорофилла. В этом случае энергия возбуждения триплетного хлорофилла и синглетного кислорода резонансным путем передается на каротиноиды, а затем рассеивается в виде тепла.
Таким образом, активный ацетил, образующийся из пировиноградной кислоты, служит исходным веществом для биосинтеза в растениях, сложных соединений к каким относятся и каротиноиды

Прорастание семян можно определить как фундаментальный процесс, начальный этап онтогенеза растений, посредством которого из одного семени формируется росток и впоследствии взрослое растение. Этот процесс влияет как на урожайность, так и ее на качество. Распространенным примером прорастания семян является образование ростка из семян покрытосеменных или голосеменных растений.
Давайте подробнее рассмотрим процесс прорастания семян, его этапы, условия и факторы необходимые для его осуществления.
Процесс прорастания семян

Полный процесс прорастания семян осуществляется в следующие фазы:
- На начальной стадии прорастания семена быстро впитывают воду, что приводит к набуханию и размягчению семенной оболочки при оптимальной температуре. Эти фазы называется водопоглощением и набуханием семян. На этом этапе запускается процесс роста путем активации ферментов. Семя активизирует свою внутреннюю физиологию и начинает дышать, вырабатывать белки и метаболизировать запасенную пищу.
- При разрыве семенной оболочки появляется корешок, образующий первичный корень. Семя начинает поглощать воду из почвы. После появления корешка и проростка, побег начинает расти вверх.
- На заключительном этапе прорастания клетки семян становится метаболически активными, удлиняются и делятся, чтобы дать начало ростку.
Условия, необходимые для прорастания семян

Вот некоторые важные требования, которые необходимы для того, чтобы семя образовало росток, а затем и растение:
- Вода: крайне необходима для прорастания семян. Некоторые семена чрезвычайно сухие и нуждаются в значительном количестве воды относительно сухого веса. Вода играет важную роль в прорастании семян. Она обеспечивает необходимую гидратацию для жизнедеятельности протоплазмы и растворенный кислород для растущего зародыша, смягчает семенную оболочку и повышает проницаемость семян. Вода помогает в разрыве семян, а также превращает питательные в растворимую форму для ее транслокации в эмбрион.
- Кислород: важный и незаменимый источник энергии для роста семян. Он необходим прорастающему семени для обмена веществ и используется как часть аэробного дыхания, пока растение не вырастить собственные зеленые листья. Кислород можно найти в порах частиц почвы, но если семя зарыто слишком глубоко, оно будет лишено этого кислорода.
- Температура: для быстрого прорастания семян требуется умеренная температура около 25-30°C. Совершенно очевидно, что различные семена нуждаются в разных оптимальных температур. Есть некоторые семена, которым необходимы либо более низкие, либо более высокие температуры от 5 до 40°C.
- Свет или тьма: это может действовать как экологический фактор. Многие семена не прорастают, пока на них не падает солнечный свет.
Процесс прорастания семян запускается при указанных выше благоприятных условиях. Семена претерпевают быстрое расширение и рост зародыша с последующим разрывом покровных слоев и появлением корешка. Появление этого корешка считается завершающей фазой процесса прорастания.
Факторы, влияющие на прорастание семян

Существует несколько основных факторов, оказывающих воздействие на прорастание семян. К ним относятся:
Внешние факторы
- Вода: недостаток или избыток воды влияет на прорастание семян.
- Температура: влияет на скорость роста, а также на метаболизм семени.
- Кислород: прорастающие семена активно дышат и высвобождают энергию, необходимую для их роста, поэтому недостаток кислорода влияет на их всхожесть.
В некоторых случаях температура ниже умеренного уровня замедляет прорастание семян и способствует росту грибов. Прорастание также может прекращается при температуре выше умеренного уровня.
Внутренние факторы
Семенной покой – это состояние, при котором семена не могут прорасти даже при благоприятных условиях. Во время семенного покоя:
Промышленное выращивание растений на питательных растворах:
2. Этап роста и развития, на котором растение испытывает наибольшую потребность в питательных веществах:
+ наиболее энергичное увеличение массы растений;
3. Элемент, максимально потребляемый растением в период интенсивного роста и образования вегетативной массы:
4. Минеральные удобрения обеспечивают прибавку урожая в среднем на, %:
5. Этап в жизни растений, на котором применяются подкормки роста:
+ усиленный рост и образование вегетативной массы;

6. Оптимальные значения почвы для большинства с.-х. растений:
7. Почему ионы и не удерживаются в почве и вымываются?
– между ними и твердыми частицами почвы наблюдается положительная адсорбция;
+ между ними и твердыми частицами почвы наблюдается отрицательная адсорбция;
– вследствие синергизма полов;
– вследствие антагонизма полов.
8. Подавляющее количество питательных веществ содержится в почве:
– нет четкой зависимости.
9. Растения поглощают питательные вещества:
– по градиенту концентраций;
10. Зависят ли коэффициенты использования элементов питания растениями от генетических свойств сорта?
– да, но только при высоком уровне урожайности;
– да, но только при низком уровне урожайности.
11. Не содержатся в составе корневых выделений:
– физиологически активные вещества.
12. Основная причина накопления нитратов в растения:
– кислая реакция почвы;
– недостаток влаги в почве;
+ избыток азота в почве;
– недостаток фосфора в почве.
13. Способность усваивать молекулярный азот из атмосферы обладают:
– все виды однолетних трав;
– все виды многолетних трав;
14. Автор теории минерального питания растений:
15. Антагонизм наблюдается между ионами:
+ кальция и калия;
– и ;
– калия и молибдена;
16. Элемент, обладающий наименьшей способностью оттекать из стареющих листьев:
17. Восходящий ток ионов осуществляется по:
– одинаково и по ксилеме, и по флоэме;
– нет четкой зависимости.
18. Зона корня, наиболее активно поглощается вещества из почвы:
– все зоны одинаково;
19. Относится к микроэлементам:
20. Для снижения кислотности почвы проводят:
21. Недостаток какого элемента вызывает хлороз растений?
22. Недостаток какого элемента резко тормозит рост растений?
Обмен веществ состоит из двух процессов
+ анаболизма и катаболизма
- анаболизма и параболизма
- параболизма и гетероболизма
- анаболизма и гетероболизма
Транспортная форма углеводов в растениях
Белки при прорастании семян
+ распадаются на аминокислоты, из которых синтезируются новые белки
- окисляются в процессе дыхания
- используются для синтеза жиров
- используются для синтеза углеводов
Жиры при прорастании семян масличных культур
- используются для синтеза белков
+ превращаются в углеводы
- превращаются в новые жиры
- непосредственно окисляются в процессе дыхания
Гидролитическое расщепление крахмала осуществляют ферменты
– совокупность всех генов
морфологические изменения растений
2. Жизненные формы растений:
– однолетние, яровые, озимые
– двулетние, многолетние, эфемеры
+ однолетние, двулетние, многолетние;
– озимые, яровые, эфемеры.
3. Типы онтогенеза у однолетников:
5. Какой подход не используется для периодизации онтогенеза:
– по фенологическим фазам;
+ по стадиям развития;
– по этапам органогенеза;
– по возрастным периодам.
Вакуоль образуется в фазе роста и развития клетки:
– старения и отмирания.
– увеличение размеров и числа клеток;
– увеличение объема клеток;
+ новообразование цитоплазмы и клеточных структур, приводящие к увеличение числа размеров клеток, тканей, органов и всего растения в целом;
– качественные изменения структуры и функции растения в целом и его отдельных частей, возникающие в процессе онтогенеза.
Определение развития растений:
– увеличение размеров и числа клеток;
– увеличение объема клеток;
+ новообразование цитоплазмы и клеточных структур, приводящие к увеличение числа размеров клеток, тканей, органов и всего растения в целом;
– качественные изменения структуры и функции растения в целом и его отдельных частей, возникающие в процессе онтогенеза.
9. Последовательность фаз роста и развития клеток:
+ эмбриональная, растяжения, дифференциации, старение и отмирание;
– дифференциации, эмбриональная, растяжения, старение и отмирание;
– эмбриональная, дифференциации, растяжения, старение и отмирание;
– растяжения, дифференциации, старение и отмирание, эмбриональная.
10. Фитогормон, контролирующий прохождение фазы растяжения роста и развития клетки:
– совместное действие трех групп фитогормонов.
11. Свет тормозит прохождения фазы роста и развития клетки:
12. Прохождении какой фазы роста и развития клеток особенно важна роль воды :
13. В какой фазе роста и развития клетки увеличивается отложение в клеточной стенке целлюлозы, лигнина, суберина:
– старения и отмирания.
14. Назовите группы фитогормонов:
– ауксины, витамины, гиббереллины, цитокинины, абсцизовая кислота;
– гиббереллины, ферменты, ауксины, этилен, абсцизовая кислота;
– абсцизовая кислота, этилен, алкалонды, витамины, ферменты;
– ауксины, гиббереллины, цитокинины, абсцизовая кислота, этилен.
15. Назовите природные фитогормоны – стимуляторы роста:
– ауксины, гиббереллины, абсцизовая кислота;
– ауксины, абсцизовая кислота, цитокинины;
– абсцизовая кислота, этилен, гиббереллины;
+ ауксины, гиббереллины, цитокинины.
16. Какой из перечисленных фитогормонов не относится к стимуляторам роста:

– ауксин (НУК);
17. Природные фитогормоны – ингибиторы роста:
– ауксины, абсцизовая кислота;
+ абсцизовая кислота, этилен;
– цитокинины, абсцизовая кислота.
18. – индолилуксисная кислота (НУК) относится к:
19. Функции этилена способствуют:
– разрастанию завязи партенокарпических плодов и образованию корней;
– ускоряет цветение короткодневных растений;
– регулирует устьичные движения является фактором физиологического покоя;
+ ингибирует рост, регулирует процессы старения, опадения листьев и генеративных органов, ускоряет созревание плодов.
20. Обработка ауксинами способствуют:
– разрастанию завязи плодов, образованию боковых корней, уменьшению опадения завязей, апикальному доминированию;
– цветению длиннодневных растений, переходу двулетников к цветению без яровизации;
– формированию почек и росту побегов, выходу из состояния покоя семян и клубней;
– созреванию плодов, смещению пола у цветков в женскую сторону.
21. Содержание какого фитогормона возрастает при действии неблагоприятных условий:
+ абсцизовой кислоты (АБК).
22. Фитогормоны, стимулирующие деление клеток, формирование почек и рост побегов, выводящие из состояния покоя клубни и семена:
23. Фитогормоны, способствующие удлинению стебля, укрупнению листьев, цветков, соцветий, ускорению цветения длиннодневных растений, переходу двулетников к цветениию без яровизации, смещению пола растений в мужскую сторону:
24. Фитогормон, накапливающий в сочных плодах в период их созревания?
25. Фитогормоны, синтез которых локализован преимущественно в верхушечных меристемах стебля и корней.
26. Фитогормоны, синтез которых локализован преимущественно в апикальной меристеме корней:
27. Фитогормоны, способствующие образованию партенокарпических плодов:
28. На каком участке большой кривой роста происходит интенсивный рост:
– начальном (лаг – период);
28 / . На логарифмическом участке большой кривой роста происходит:
– зависимость роста и развития одних органов, тканей или частей растения от других, их взаимное влияние;
– вегетативное размножение растений;
+ восстановление поврежденных или утраченных частей;
– неравноценность противоположных полюсов клетки, органа, целого растения.
30. Свойство, связанное с неравномерностью противоположных полюсов клетки, органа, целого растения:
31. Какие приборы используются для измерения скорости роста:
32. Прямое действие света на рост растений?
– рост стебля усиливается;
+ рост стебля уменьшается;
– листья вырастают крупнее;
– листья приобретают темно-зеленую окраску.
33. Какие лучи способствуют вытягиванию, ветвлению растений?
34. Какие растения называются этиолированными?
+ выращенные в темноте и лишенные хлорофилла;
– обработанные органическими растворителями;
– выращенные при недостатке элементов питания;
– выращенные на синем свету.
35. Гормональный оптимум – это температуры:
+ благоприятные для роста растений;
– благоприятные для развития растений;
– не оказывающие существенного влияния на рост и развитие растений;
– благоприятные и для роста, и для развития растений;
36. Как называется реакция растений на периодическую смену повышенных и пониженных температур, выражающаяся в изменения процессов роста и развития?
37. Какой тип роста характерен для корней?
38. Благоприятные для роста условия при влажности почвы, % НВ:
39. Для улучшения лежкости плодов и овощей при их хранения используется:
40. Как называются биологические ритмы с периодичностью около 24 часов?
41. Ростовые движения растений в ответ на односторонне действующий раздражитель называются:
42. Какой гормон играет решающую роль в явлении фототропизма?
43. Отрицательный геотропизм характерен для:
44. Настин – это движения, которые возникают у растений в ответ на:
– односторонне действующий фактор среды;
+ на диффузное (равномерное со всех сторон) действие фактора;
– при изменении градиента концентрации питательного раствора;
– нет четкой зависимости.
45. Изменения положения органов растений, вызываемых прикосновением, называются:
46. Укажите правильную последовательность этапов онтогенеза:
– ювениальный, эмбриональный, размножение, зрелость, старение;
– эмбриональный, ювениальный, размножение, зрелость, старение;
+ эмбриональный, ювениальный, зрелость, размножение, старение;
– эмбриональный, размножение, ювениальный, зрелость, старение;
47. Какие растения в первый год жизни образуют вегетативные органы, а во второй год цветут и плодоносят?
48. Яровизация – это стимуляция цветения путем воздействия на растения:
– низких отрицательных температур;
+ низких положительных температур;
температур около 0 0 ;
49. Для каких растений яровизация является обязательным условием их нормального развития?
50. В состоянии покоя растения характеризуется:
+ отсутствием видимого роста;
– высокой интенсивностью дыхания;
– высокой скоростью превращения веществ;
– низкой устойчивостью к неблагоприятным факторам.
51. Какой вид покоя наблюдается при неблагоприятных внешних условиях:
– глубокий (органический, физиологический);
52. Стратификация как прием обработки семян:
– стимулирует цветение растений;
– продлевает покой семян;
+ способствует прорастанию семян.
53. Условия стратификации:
+ влажный песок и пониженная температура;
– механическое повреждение оболочки семян;
– повышенная температура (20 – 25 0 );
54. Фотопериодизм – это способность растений переходить к цветению в зависимости
– от перепада ночных и дневных температур;
– от смены дня и ночи;
+ при определенном соотношении длины темного и светлого периода суток;
– при непрерывном освещении.
55. Зацветание растений короткого дня происходит при фотопериоде длиной (часов):
56. Оптимальная длина фотопериода для растений длинного дня (часов):
57. Органом, воспринимающим фотопериодическую реакцию, является:
58. Задержать старение однолетних растений можно:
– удалением отдельных органов;
+ усилением азотного питания;
59. Какие вещества применяются для предотвращения полегания растений?
60. Какой фактор не способствует полеганию растений?
– высокий уровень азотного питания;
– высокая влажность почвы;
61. Вещества, применяемые для ускорения опадения листьев:
62. Вещества, применяемые для ускорения обезвоживания и засыхания листьев:
63. Прием, ускоряющий прорастания семян с плотной оболочкой:
64. Признак, объединяющий вынужденный и глубокий покой растений:
+ отсутствие видимого роста;
– наличие неблагоприятных условий;
– отсутствие фактора, необходимого для ростовых процессов;
– необходимость определенных физиолого-биохимических измерений.
65. Движения, вызванные неравномерным освещением разных сторон органа:
66. Движения, вызванные односторонним влиянием силы тяжести:
67. Движения, вызванные толчками:
68. Активной формой фитохрома является:

– ;

+ ;

– ;

– .
69. Фотоморфогенез, т.е. зависимые от света процессы роста и дифференцировки растений, определяющие их форму и структуру контролируются:
Фотосинтез является единственным механизмом получения энергии для биосферы (за исключением процессов, происходящих у некоторых хемосинтезирующих бактерий, которые получают энергию от окисления неорганических субстратов). Фотосинтез включает в себя окислительно-восстановительные реакции. В общем, в процессе окисления молекулы воды высвобождаются электроны с выделением кислорода и восстановлением диоксида углерода с образованием углеводов (Salisbury и Ross, 1994).

Фотосинтез важен для человека, среди прочего, потому что через него производится пища и кислород (Barcelo и др., 1983). Кроме того, он прямо или косвенно кормит почти весь живой мир на планете (Campbell и Reece, 2005).
Первые организмы-фотосинтезаторы, вероятно, появились 3 миллиарда или 3,5 миллиардов лет назад. По мере их распространения они устанавливали взаимозависимости и отношения, которые не только трансформировали их, но и изменили внешний облик планеты. Древние организмы жили в среде без свободного кислорода в атмосфере, в состав которой входил большой процент водорода. Кислород, даже был токсичным для этих примитивных организмов, так же, как в настоящее время для анаэробов. Энергия приобреталась такими процессами, как ферментация или гликолиз, что должно было привести к постепенному накоплению углекислого газа в ранней атмосфере. Если бы живые существа использовали сразу солнечную энергию, используя углекислый газ и выделяя кислород, они, безусловно, более эффективно использовали бы ресурсы своей окружающей среды, отметив веху в истории и эволюции жизни на планете. Условием фотосинтеза является поглощение фотонов, квантовых частиц определенными пигментами (Curtis и Barnes, 2001).
Фотосинтез представляет собой эндергонический процесс, потому что необходимо участие энергии солнечного излучения, чтобы начать цепные реакции, которые приводят к образованию органических соединений, которые накапливает организм.
Общая реакция фотосинтеза:
(CH2O)n- это сокращенная форма представления крахмала или других углеводов по эмпирической формуле. Крахмал является наиболее распространенным продуктом фотосинтеза. Фотосинтез зависит от различных факторов окружающей среды, таких как интенсивность и качество освещения, влажность воздуха, температура, наличие воды, минеральных веществ и углекислого газа. Фотосинтез одного листа опирается на более, чем 50 индивидуальных реакций, каждая из которых зависят от параметров окружающей среды (Cogua, 2011).
Способность растений компенсировать воздействия окружающей среды зависит от их производительности и выживания в конкретной среде. Скорость фотосинтеза может измениться в течение следующих десятилетий в зависимости от приспособительных реакций к изменению уровней CO2, начиная от прямых реакций собственно на CO2, вплоть до косвенных реакций в связи с изменением температуры и водного режима, которые могут произойти в будущем (Cogua, 2011).
Хлоропласты
Клеточными органеллами, в которых происходит фотосинтез, являются хлоропласты.
Пигменты фотосинтеза
Чтобы световая энергия могла быть использована живыми существами, она должна быть поглощена фотосинтезирующими пигментами. Эти пигменты представляют собой вещества, способные поглощать свет на определенных длинах волн, например, хлорофилл – пигмент, который придает растениям характерный зеленый цвет, поглощает свет на длинах волн, соответствующих фиолетовому, синему и красному цвету, отражая зеленый цвет (Azcon и Talon, 1993).
Хлорофилл
Структура молекулы хлорофилла
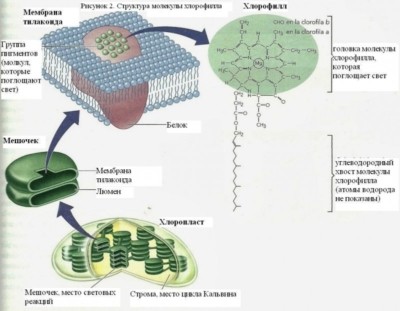
Хлорофилл является тетрапиррольным соединением, т.е. состоит из четырех пиррольных колец, соединенных метильными мостиками, которые образуют порфирин. Тетрапиррол является основной структурой порфирина, который, как и в хлорофилле, является частью гемоглобина и цитохрома.

Хлорофиллы a и b
Как упоминалось выше, хлорофилл a – это пигмент, непосредственно связанный с превращением световой энергии в химическую. Тем не менее, существует еще один тип хлорофилла, хлорофилл b, и пигменты, называемые каротиноидами, которые бывают красными, оранжевыми или желтыми, и которые в зеленых листьях маскируются обилием хлорофиллов. Эти дополнительные пигменты хлорофилла позволяют растениям поглощать более широкий диапазон спектра света, доступного для фотосинтеза, действуя в качестве рецепторов, которые передают энергию. Также существует хлорофилл с, найденный в бурых водорослях. В красных водорослях найден хлорофилл d. Наконец, существует тип хлорофилла (бактериохлорофилл), который является пигментом фототрофных бактерий (Cogua, 2011).
Каротиноиды
Желтые и красные пигменты, известные как каротиноиды, имеют систему сопряженных двойных связей, образованных атомами углерода, являются соединениями, нерастворимыми в воде, но растворимы в жировых растворителях. Они разделены на ненасыщенные углеводороды или каротины и кислородсодержащие производные, называемые ксантофиллами. В растениях каротиноид называется бета-каротином. Это пигменты красного, оранжевого или желтого цвета (Curtis и Barnes, 2001). Каротиноиды, участвующие в процессе фотосинтеза, называются первичными, в отличие от тех, которые находятся в цветах и плодах, формируя хромопласты, и в гетеротрофах, таких как бактерии, дрожжи и грибы (Cogua, 2011).
Организация пигментов в тилакоидах
Как отмечено выше, структурные единицы фотосинтеза у эукариот представляют собой хлоропласты, расположенные преимущественно в листьях. Внутренние ткани каждого листа покрыты эпидермальными клетками, покрытыми кутикулой, восковым слоем. Кислород и углекислый газ проникают в листья через специальные поры, называемые устьицами. Высокий процент фотосинтеза происходит в клетках палисадной паренхимы, которые расположены под эпидермисом и образуют мезофилл. Хлоропласты ориентированы внутри клеток, чтобы облегчить поглощение света тилакоидными мембранами (Curtis and Barnes, 2001).
Стадии фотосинтеза
Английский физиолог Ф. Блэкмэн (F. F. Blackman) после проведения различных экспериментов предположил, что в процессе фотосинтеза существует, по крайней мере, два ограничивающих фактора: интенсивность света и температура. Поэтому существует группа реакций, которые зависят от света, но не зависят от температуры, как видно из общей схемы фотосинтеза (Рисунок 3) (Curtis and Barnes, 2001).
Общая схема фотосинтеза

Поглощение света
Световые реакции можно резюмировать следующим образом. Светопоглощающие пигменты делятся на две группы: те, которые поглощают и передают энергию к реакционному центру, и те, которые образуют этот реакционный центр, представляющий собой определенный тип молекул хлорофилла (хлорофилл a P680 и Р700) и осуществляют фотохимическую реакцию.
Для обеспечения фотосинтеза требуется сотрудничество двух типов фотосистем, называемых I и II (PS I и PS II). В PS II вода используется в качестве донора электронов, выделяя кислород в качестве побочного продукта. В PS I восстанавливается терминальный акцептор, отдавая электрон ферредоксину, чтобы с его помощью восстановить НАДФ (NADP), которая, наконец, используется для превращения CO2 в углеводы (Melgarejo, 2010).
Фотосистемы I и II работают вместе, хотя в то же время фотосистема I может работать независимо, как это описано на рисунке.
В данной модели энергия света достигает фотосистемы II, где она захватывается молекулой хлорофилла a P680. Электрон молекулы Р680 высвобождается на более высоком энергетическом уровне, из которого он передается к молекуле первичного акцептора электронов. Электрон проходит вниз на фотосистему I вдоль цепи переноса электронов. В результате потока электронов вдоль транспортной цепи устанавливается протонный градиент по всей тилакоидной мембране; потенциальная энергия этого электрохимического градиента используется для формирования АТФ (ATP) из АДФ (ADP) – хемиосмотического процесса, аналогичного процессу у митохондрий. Этот процесс известен как фотофосфорилирование (Curtis и Barnes, 2001).
Одновременно выделяются три других действия:
1. Молекула хлорофилла P680, теряя свой электрон, ищет замену. Она находит ее в молекуле воды, которая, будучи прикрепленной к молекуле, содержащей марганец, лишается электрона, а затем распадается на протоны и газообразный кислород.
2. Дополнительно световая энергия захватывается в реактивной молекуле хлорофилла Р700 фотосистемы I. Молекула окисляется, и электрон высвобождается в первичный акцептор электронов, из которого он перемещается вниз по направлению к НАДФ+ (NADP+).
3. Электрон, удаленный из молекулы P700 фотосистемы I, замещается электроном, который перемещался вниз от первичного акцептора электронов фотосистемы II. Таким образом, на свету происходит непрерывный поток электронов от воды к фотосистеме II, к фотосистеме I, к НАДФ+ (CURTIS, Helena, BARNES, Sue, 2001). Энергия, восстановленная на этих этапах, представлена молекулой АТФ (ATP) (образование которой высвобождает молекулу воды) и НАДФH (NADPH), которые затем преобразуются в основные источники энергии для восстановления углекислого газа. Для генерации одной молекулы НАДФН необходимо поглотить 4 протона: 2 фотосистемой II и 2 фотосистемой I (Curtis и Barnes, 2001).
Восстановление и фиксация CO2
На второй стадии фотосинтеза энергия используется для восстановления углерода. Углерод доступен для фотосинтезирующих клеток в форме диоксида углерода. У растений двуокись углерода достигает фотосинтезирующих клеток через специализированные отверстия листьев и зеленых стеблей, называемые устьицами (Curtis и Barnes 2001).
На этой второй стадии фотосинтеза, АТФ и НАДФН, сформированные на первой стадии, используются для восстановления углерода из диоксида углерода до простого сахара. Затем химическая энергия, временно хранящаяся в молекулах АТФ и НАДФН, передается соответствующим молекулам для транспортировки и хранения энергии в клетках водорослей или органов растений. Результатом этого процесса является формирование углеродного скелета, из которого затем могут быть построены и другие органические молекулы (Cogua, 2011).
Реакции фиксации углерода проходят в условиях, которые не требуют света, но некоторые ферменты регулируются им и производятся в строме посредством цикла Кальвина (Рисунок 5). НАДФ и АТФ, образованные в реакциях, которые поглощают световую энергию, используются для восстановления углекислого газа. Цикл производит глицеральдегид фосфат, из которого может быть сформирована глюкоза и другие органические соединения (Cogua, 2011).
Цикл Кальвина

Это преобразование происходит в строме хлоропластов. CO2 поступает в хлоропласт, проходя через клеточные мембраны органеллы по определенным каналам. Этот газ поступает в тело растения, благодаря устьичному отверстию, которое в то же время облегчает выделение водяного пара в процессе, известном как транспирация (Cogua, 2011). Цикл Кальвина представляет собой центральный путь восстановления СО2 до органических молекул. Он начинается, когда фермент рибулоза 1,5 бифосфат карбоксилаза оксигеназа (Rubisco) выполняет карбоксилирование СО2 и дает начало двум молекулам из трех атомов углерода (Melgarejo, 2010). Чтобы выяснить это, было проведено несколько экспериментов, которые были успешными, благодаря использованию радиоизотопов в зеленых водорослях Chlorella pyrenoidosa и Scenedesmus obliquus. После получения 14СО2 в течение 60 секунд в культурах вышеупомянутых водорослей появляется несколько меченых соединений; фосфаты сахаров, органические кислоты и аминокислоты. Сокращая время применения С14О2 шаг за шагом, удалось восстановить образованные меченые продукты. Менее чем за 2 секунды появляется только одно меченое соединение, 3-фосфоглицериновая кислота (3-PGA), состоящая из трех атомов углерода. Таким образом, первая стадия восстановления СО2 состоит в карбоксилировании, в ходе которого СО2 включается в молекулу акцептора, имеющую 5 атомов углерода, рибулозо-1,5-дифосфат, расщепление которого приводит к образованию двух молекул 3-PGA. Механизм восстановления СО2 является циклическим процессом из нескольких частичных ферментативных стадий (Мельгарехо, 2010).
Метаболизм Хэтча/Слэка

Существуют и другие адаптивные процессы у различных типов растений, например, некоторые растения присоединяют диоксид углерода к первому соединению, известному как фосфоенолпировиноградная кислота или фосфоенолпируват (РЕР) с образованием соединения из четырех атомов углерода, щавелевоуксусной кислоты (промежуточное соединение в цикле Кребса). Среди других растений, которые имеют этот тип метаболизма, кукуруза и сорго. У них диоксид углерода, включенный в щавелевоуксусную кислоту, окончательно переносится в RuBP и входит в цикл Кальвина, но только после прохождения ряда реакций, которые транспортируют его в более глубокие участки внутри листа. Эти реакции протекают в клетках мезофилла, где хлоропласты характеризуются наличием обширной сети тилакоидов, организованных в хорошо развитые граны. Яблочная или аспарагиновая кислота, в зависимости от вида, транспортируются к клеткам обкладки сосудистого пучка, благодаря сети плазмодесм. Хлоропласты этих клеток, которые образуют плотные оболочки вокруг сосудистых пучков листа, имеют неразвитые граны и часто содержат граны крахмала. Растения, использующие этот путь, или путь Хэтча-Слэка, обычно называют C4-растениями, благодаря 4-углеродному соединению, которое служит в качестве акцептора СО2 (Cogua, 2011).
Кислотный метаболизм Толстянковых или растения MAC (CAM)

Существуют некоторые виды растений сухого климата, которые имеют толстые листы, с низким соотношением поверхности к объему, толстой кутикулой и низкой скоростью транспирации. Такие растения называются суккулентами. Как правило, фотосинтезирующие клетки листьев или стебля у них представлены в губчатом мезофилле. У некоторых суккулентов метаболизм CO2 необычен, первоначально исследован у членов семейства Толстянковых, поэтому его называют Кислотный метаболизм Толстянковых (Crasulaceae Acid Metabolism (CAM или MAC) по-английски).
Новый фотосинтетический путь, упомянутый выше, был найден у некоторых растений суккулентов или полусуккулентов из родов Бриофи́ллум, Каланхоэ, Седум, Клейния, Толстянка, Опунция (Salisbury и Ross, 1994). MAC или CAM растения поглощают диоксид углерода в течение ночи, фиксируя в органических кислотах, которые подкисляют среду и высвобождают его в течение дня, чтобы сразу же использовать в цикле Кальвина. Этот тип метаболизма уменьшает потери воды, связанные с открытием устьиц. (Cogua, 2011).
Факторы окружающей среды и фотосинтез
Подсчитано, что количество углерода, который фиксируется каждый год на планете, колеблется от 70000 до 120000 млн. тонн или эквивалентно около 170 – 290 гигатоннам сухого вещества, с минимальной формулой 0. Эта огромная производительность имеет место, несмотря на низкую концентрацию в атмосфере. Более 13% атмосферного углерода используется каждый год в процессе фотосинтеза, и почти равное количество обменивается при растворении в океанах. На фотосинтез оказывают влияние такие факторы, как вода, концентрация СО2, свет, питательные вещества и температура, а также возраст и генетика растений (Salisbury и Ross, 1994).
CO2
Количество СО2 является фактором, определяющим производительность, тем не менее некоторые реакции фотосинтеза могут быть осуществлены в его отсутствие. Однако, без этого газа не будет синтеза углеводов. Концентрация CO2 в атмосфере не является оптимальной для фотосинтеза, в сельскохозяйственной практике используется искусственное добавление газообразного СО2 в условиях постоянного освещения, для увеличения фотосинтетической скорости и урожайности при производстве биологического материала (Salisbury и Ross, 1994). CO2, как известно, производит немедленное увеличение скорости фотосинтеза, особенно у C3-растений. Однако, когда растения растут непрерывно с повышенным уровнем СО2, происходят биохимические изменения, которые снижают фотосинтетическую способность листьев, поэтому большой начальный рост фотосинтеза при высокой концентрации СО2 обычно не сохраняется таким же высоким, когда проходят несколько недель или месяцев. Это явление известно как акклиматизация фотосинтеза (Cogua, 2011).
Вода
Внеклеточно у высших растений вода транспортирует такие материалы, как соли, которые перемещаются от корней ко всем частям растения. Помимо того, что она является сырьем в процессе фотосинтеза, вода способствует в качестве реагента нескольким метаболическим реакциям. Ионы, образующие молекулу воды (Н) и (ОН) собираются и рекомбинируются в новые молекулы воды (Cogua, 2011).
Свет
Нет света, нет фотосинтеза, растения нуждаются в свете с точки зрения интенсивности и качества излучения. С увеличением интенсивности света увеличивается фотосинтетическая интенсивность вначале по линейной зависимости, а затем плавно уменьшается и, наконец, достигает постоянного значения, т.е. фотосинтетическая способность насыщается светом. Это значение насыщения достигается различными видами с разной скоростью. У светолюбивых растений это происходит после достижения высокой интенсивности излучения, а у тенелюбивых растений это насыщение достигается быстро, то есть требуется низкая интенсивность света (Cogua, 2011).
Температура
Зависимость фотосинтеза от факторов света и температуры позволяет сделать вывод, что фотосинтез не является непрерывным процессом, а состоит из множества фотохимических реакций, которые зависят от света и серии ферментативных реакций, зависящих от температуры. Последние очевидны в состоянии насыщения света, точке, в которой повышение температуры увеличивает фотосинтетическую интенсивность (Cogua, 2011). Транспорт, промышленность, вырубка лесов, сельское хозяйство и другие виды деятельности человека вызывают увеличение концентрации CO2 и других газов в атмосфере, таких как метан. Скопление этих газов имеет тенденцию к нагреву атмосферы, что может привести к региональным или глобальным изменениям, которые повлияют на такие параметры, как температура, осадки, влажность почвы и уровень моря, все они связаны с фотосинтезом и, следовательно, с самой жизнью на планете. (Cogua, 2011).
Инга Костенко, Mivena,Украина
Анна Устименко, Клуб Sirius Agro Plant
Читайте также: