
Пути повышения продуктивности фотосинтеза в посевах
Обновлено: 20.05.2024
СЕЛЕКЦИЯ / КУЛЬТУРА / СОРТ / ФОТОСИНТЕЗ / ЭНЕРГЕТИЧЕСКИЙ ПОТЕНЦИАЛ / ГЕНОФОНД / УРОЖАЙНОСТЬ / КАЧЕСТВО / СТАБИЛЬНОСТЬ / BREEDING / CROPS / VARIETIES / PHOTOSYNTHESIS / PHOTOENERGY POTENTIAL / EFFICIENCY / YIELD CAPACITY / QUALITY / SUSTAINABILITY
Аннотация научной статьи по сельскому хозяйству, лесному хозяйству, рыбному хозяйству, автор научной работы — Амелин А.В.
В статье представлены результаты многолетних собственных научных исследований (1983-2015 гг.) и анализ литературных экспериментальных данных отечественных и зарубежных ученых по проблеме повышения активности и эффективности фотосинтеза культурных растений средствами селекции . Сделано заключение, что физиологической основой селекции сельскохозяйственных культур служит система регуляторных механизмов, имеющих компенсаторный характер, позволяющих за счет разной морфофизиологической организации продукционного процесса растений достигать одного и того же результата повышения продуктивности в различных природно-климатических условиях выращивания. При этом, потенциальные возможности фотоэнергетического процесса растений в результате селекции не претерпевают больших изменений и в целом балансируют на определенном для биологического вида уровне, что сдерживает дальнейший прогресс селекции . Для изменения ситуации рекомендуется формы с повышенной интенсивностью фотосинтеза , фотохимической активностью хлоропластов и энергетической эффективности электронно-транспортной цепи брать на учет и использовать в селекции как исходный перспективный материал. У гороха, гречихи, сои, чечевицы и кормовых бобов установлена значительная генотипическая вариация значений интенсивности фотосинтеза , реакции Хилла и фотофосфорилирования, квантового выхода и фотохимического тушения флуоресценции хлорофилла
Похожие темы научных работ по сельскому хозяйству, лесному хозяйству, рыбному хозяйству , автор научной работы — Амелин А.В.
Селекция на повышение фотоэнергетического потенциала растений и эффективности его использования, как стратегическая задача в обеспечении импортозамещения и продовольственной безопасности России
Влияние температуры, увлажнения и фазы роста на интенсивность фотосинтеза листочков и прилистников растений гороха посевного
Морфофизиологические параметры перспективного зернового сорта гороха полевого для центральных регионов России
Влияние интенсивности света на активность газообмена листьев и прилистников у белоцветковых сортов гороха
INCREASE OF ACTIVITY AND EFFICIENCY OF PHOTOSYNTHESIS OF CULTIVATED PLANTS BY BREEDING
The article presents the results of years of their research (1983 to 2015) and the analysis of published experimental data of domestic and foreign scientists on the problem of increasing the activity and efficiency of photosynthesis of cultivated plants by means of breeding . It is concluded that the physiological basis of crop breeding is the system of regulatory mechanisms that are compensatory in nature, allowing for different morph physiological organization of plant production process to achieve the same result the increase of productivity in different climatic growing conditions. At the same time, the potential of photovoltaic process plants as a result of selection do not undergo large changes and overall balance on a specific to a species level, which hinders the progress of breeding . To change the situation it is recommended that forms with increased intensity of photosynthesis , photochemical activity of chloroplasts and the energy efficiency of the electron transport chain to take account of and use in breeding as a source of promising material. Pea, buckwheat, soybeans, lentils and broad beans showed large genotypic variation in the intensity of photosynthesis , the hill reaction and photophosphorylation, quantum yield and photochemical quenching of chlorophyll fluorescence
УДК 631.524.8:338.439.01.001.25 (471+571)
ПОВЫШЕНИЕ АКТИВНОСТИ И ЭФФЕКТИВНОСТИ ФОТОСИНТЕЗА КУЛЬТУРНЫХ РАСТЕНИЙ С ПОМОЩЬЮ СЕЛЕКЦИИ
В статье представлены результаты многолетних собственных научных исследований (1983-2015 гг.) и анализ литературных экспериментальных данных отечественных и зарубежных ученых по проблеме повышения активности и эффективности фотосинтеза культурных растений средствами селекции. Сделано заключение, что физиологической основой селекции сельскохозяйственных культур служит система регуляторных механизмов, имеющих компенсаторный характер, позволяющих за счет разной морфофизиологической организации продукционного процесса растений достигать одного и того же результата -повышения продуктивности в различных природно-климатических условиях выращивания. При этом, потенциальные возможности фотоэнергетического процесса растений в результате селекции не претерпевают больших изменений и в целом балансируют на определенном для биологического вида уровне, что сдерживает дальнейший прогресс селекции. Для изменения ситуации рекомендуется формы с повышенной интенсивностью фотосинтеза, фотохимической активностью хлоропластов и энергетической эффективности электронно-транспортной цепи брать на учет и использовать в селекции как исходный перспективный материал. У гороха, гречихи, сои, чечевицы и кормовых бобов установлена значительная генотипическая вариация значений интенсивности фотосинтеза, реакции Хилла и фотофосфорилирования, квантового выхода и фотохимического тушения флуоресценции хлорофилла.
Ключевые слова: селекция, культура, сорт, фотосинтез, энергетический потенциал, генофонд, урожайность, качество, стабильность.
В настоящее время назрела необходимость использования качественно новых подходов к производству, в котором центральное место должна занять гармонизация отношений человека с природой, экономикой и экологией, так как современное растениеводство, наряду с большими достижениями, характеризуется и ярко выраженными тенденциями ухудшения эколого-биологических, агротехнологических и экономических ее факторов [1]. В решении этой проблемы весьма важная роль отводится селекции, которая добивается повышения урожайности и качества продукции за счет использования наследственного потенциала сельскохозяйственных культур. В последнее время вклад сорта в формирование урожая в развитых странах мира стал превышать 60 % [2].
Однако, новые районированные сорта отвечают не всем современным требованиям экологически чистого и энергетически эффективного производства. Они формируют высокий и стабильный урожай, как правило, в благоприятных погодных условиях и за счет использования высоких доз удобрений и большого количества химических средств защиты, так как многие из них в сильной степени поражаются болезнями и повреждаются вредителями
Изменить сложившуюся ситуацию традиционными методами селекции весьма проблематично, ввиду того, что они во многом исчерпали свои возможности [5, 6]. Морфологические признаки растений и элементы структуры урожая, по которым в основном ведется искусственный отбор, в значительной степени оптимизированы для большинства сельскохозяйственных культур в производственных зонах их преимущественного возделывания [6]. Поэтому решить стоящую проблему предлагается принципиально иным способом - посредством повышения эффективности использования солнечной радиации через
управление фотосинтезом, что позволит создать не только эффективное, но и адаптивное производство [7, 8].
К сожалению, селекция в данном направлении не осуществлялась, ни в бывшем Советском Союзе, ни за рубежом [9].
Учитывая это, впервые в России нами предпринимается попытка решить стоящую проблему за счет эффективного использования селекцией возобновляемого природного источника энергии фотосинтеза, где скрыты огромные неиспользованные резервы. Научно-методическая работа в этом направлении осуществляется с 1983 года, а целенаправленная селекционная работа с 2009 года по совместной программе с селекционерами ВНИИ зернобобовых и крупяных культур, а с 2015 года в работе задействованы и селекционеры Белгородского ГАУ им. В.А. Горина.
Опытный материал выращивался в селекционном севообороте на делянках площадью 7,5-10 м , в 4-х кратной повторности. Размещение делянок - рендомизированное.
Фотовосстановительная активность изолированных хлоропластов (ФВАХ) определялась в двух повторениях по восстановленному на свету феррицианиду калия за единицу времени. Учет феррицианида проводили в молях в расчете на моль хлорофилла в минуту (моль К4Ее(СК)б /моль Хл. мин.) с помощью спектрофотометра марки СФ-18М. Исследования осуществляли на протяжении всей вегетации: через каждые 7 дней отбирали средние пробы из третьего и четвертого листьев сверху, а с фазы образования бобов опытным материалом служили прилистники, усики, стебель, черешки, створки бобов. Выделение изолированных хлоропластов, приготовление опытных сред и проведение реакций выполняли с учетом методических рекомендаций, разработанных Могилевой Г.А. и др. 1978. Для получения суспензии изолированных хлоропластов использовали центрифугу РС-6, с автоматическим управлением частоты вращения барабана и температурного режима.
Интенсивность фотосинтеза оценивалась в режиме реального времени на интактных растениях с помощью портативного газоанализатора Li - 6400 американской фирмы Ш-СОК
Обобщение полученного экспериментального материала по всем проведенным многолетним исследованиям и имеющихся по этой проблеме литературных сведений позволило заключить, что физиологической основой селекции сельскохозяйственных культур служит адаптивная система регуляторных механизмов, имеющих компенсаторный характер, позволяющих за счет разной морфофизиологической организации продукционного процесса растений достигать одного и того же результата - повышения продуктивности в различных природно-климатических условиях выращивания.
К примеру, у растений гороха в результате длительного искусственного отбора на высокую семенную продуктивность фотоассимиляционная поверхность листьев, содержание хлорофилла и продолжительность их функционирования уменьшились на 30. 40 %, но при этом на такую же величину соответственно повысились удельная поверхностная плотность листьев (УПП), чистая продуктивность фотосинтеза (ЧПФ) и ФВАХ (рис.1).
Выявлена четко выраженная причинно-следственная связь, чем меньше листовая поверхность, тем плотнее и толще листовые пластинки и прилистники (г=+0,98), за счет развития клеток губчатой паренхимы (г=+0,94), тем активнее они фотосинтезируют (г=+0,70) и эффективнее обеспечивают семена фотоассимилянтами по известному физиологическому принципу дублирования, сформулированному Бартковым Б.И. и Зверевой Е.Г. (1974): метаболиты из одного листа одновременно поступают в бобы разных продуктивных узлов, каждый из которых обслуживается не одним, а многими листьями, но определяющую роль, по данным Pate Y.S. и Flinn A.M. (1977), выполняют ближе расположенные.
Биологический смысл этих взаимосвязей заключается в том, что повышение удельной поверхностной плотности и толщины листьев, при уменьшении их ассимиляционной поверхности, обеспечивает более эффективное улавливание квантов света и их использование в фотосинтетической деятельности растений (ФВАХ, ЧПФ), продолжительность активного периода которой у сортообразцов гороха во многом определяется развитием губчатой паренхимы, служащей местом временного депонирования запасных веществ (крахмала), когда спрос на них со стороны основных потребителей (семян) не большой (рис. 2).
Рис. 2. Поперечные срезы листовых пластинок у современного (а) и старого (б) сортов гороха
зернового использования При редукции же у растений листочков и прилистников (генотипы и
происходит усиление фотосинтетической функции и их усиков, черешков, стебля, створок
бобов, которые способны осуществлять первичные процессы фотосинтеза с такой же активностью, как и основные органы (рис. 3).
Posted in Биология Tags: Повышения продуктивности растений, Фотосинтез
Каждый год благодаря фотосинтезу на Земле образуется более 250 млрд тонн органических веществ (в сухом весе). Энергетический потенциал этой массы превышает более чем в 20 раз энергию используемых полезных ископаемых. При этом в атмосферу выделяется приблизительно 200 млрд тонн свободного кислорода. Хотя эффективность фотосинтеза довольно высока, лишь только 0,1-1% энергии падающего на Землю солнечного света запасается в виде органических соединений.
В первую очередь эти потери связаны с неполным поглощением света, ограничениями на биохимическом и физиологическом уровнях, также существуют энергозатраты на обеспечение жизнедеятельности растений. Поэтому для повышения продуктивности фотосинтеза используют целый ряд агротехнических мероприятий.
Во-первых, растениям необходимо обеспечить оптимальный световой режим, который можно регулировать определенной густотой посевов, правильными сроками посадки, искусственным освещением в теплицах и т.п., также важно выдержать оптимальный температурный режим. Важную роль играет содержание влаги в почве, которое можно регулировать с помощью искусственного полива или, наоборот, осушения. Кроме того, сложно получить высокие урожаи без внесения в почву достаточного количества органических и минеральных удобрений, а в теплицах желательно следить за содержанием углекислого газа в воздухе.
Огромное значение в повышении продуктивности сельскохозяйственных растений играет создание новых сортов с повышенной эффективностью фотосинтеза, устойчивых к болезням и вредителям, засухе и морозам, способных расти на засоленных и бедных почвах и т.д. В последнее десятилетие для выведения новых сортов растений все чаще и чаще применяют методы генетической и клеточной инженерии. Например, в клетки картофеля введены гены, которые делают растения ядовитыми для личинок колорадского жука, поэтому картофельные поля уже не нужно обрабатывать ядохимикатами. Это приводит не только к существенному уменьшению затрат, но и предохраняет от загрязнения окружающую среду. Уже созданы устойчивые к заболеваниям сорта картофеля, декоративных и ягодных культур, древесные породы и культурные растения, не чувствительные к гербицидам – веществам, используемым для борьбы с сорняками.
Ученые предпринимают попытки вывести новые растения, к примеру злаки, которые будут способны фиксировать атмосферный азот. В начале ХХI века в США, Китае и ряде других стран новые сорта растений, созданных генноинженерным путем и содержащим гены из других организмов (так называемые трансгенные растения), занимают уже миллионы гектаров сельскохозяйственных угодий.
От себя добавлю, что предпочитаю покупать продукты, которых не коснулась геннетическая инженерия.

Фотосинтез и урожай взаимосвязаны. Первичное образование органического вещества происходит в процессе фотосинтеза, но судить о конечном накоплении органической массы по интенсивности фотосинтеза нельзя, так как далеко не все продукты фотосинтеза сохраняются в теле растения. Образование органического вещества происходит в процессе фотосинтеза
Общее количество накопленного за определенный промежуток времени органического вещества зависит не только от фотосинтеза, но и от процесса дыхания. Процесс дыхания в растении идет непрерывно и осуществляется за счет органических веществ, образованных в процессе фотосинтеза.
Продуктивность фотосинтеза
Накопленное органическое вещество представляет собой разность между веществом, выработанным в процессе фотосинтеза, и веществом, расходуемым на дыхание. Суточный привес сухого вещества, отнесенный к единице площади растения, называют продуктивностью фотосинтеза.
Таким образом, чем больше продуктивность фотосинтеза, тем выше урожай растений. Продуктивность фотосинтеза (Ф) легко рассчитать по следующей формуле: Ф = (В2-В1)/[1/2(Л1+Л2)Т] (г/м 2 ·сутки), где: В2-В1 — увеличение сухой массы урожая растений за Т дней при весе их в начале учетного промежутка В1, а в конце — В2, 1/2(Л1+Л2)—средняя площадь листьев тех же растений за Т дней при размере ее в начале учетного промежутка Л1 кв. м, в конце — Л2 кв. м.
Показатель чистой продуктивности представляет собой результат образования органических веществ в процессе фотосинтеза и усвоения из почвы минеральных веществ за вычетом потерь веществ на дыхание и отмирание органов.
Продуктивность фотосинтеза растений обычно составляет 4— 6 г в сутки на 1 кв. м листьев, но может быть и больше — 8—10 и даже 12—15 г. Поэтому основная задача — увеличивать продуктивность фотосинтеза и сделать ее устойчивой.
Поэтому при выращивании растений необходимо это учитывать и принимать все меры, чтобы листья максимально поглощали падающую на них энергию солнечного света и использовали ее с наибольшим коэффициентом полезного действия. Для этого нужно увеличивать листовую площадь растений, правильно располагая растения на участках.

Взаимосвязь между интенсивностью фотосинтеза и величиной урожая очень сложна и только изредка между ними существует прямая зависимость. В то же время интенсивность фотосинтеза— основа для получения высоких урожаев. Интенсивность фотосинтеза— основа для получения высоких урожаев
Поэтому все мероприятия, которые увеличивают фотосинтез, — правильные схема посева и направление рядков, улучшение водоснабжения, дождевание и полив в районах орошения, подкормка углекислотой, правильное применение удобрений — должны привести к увеличению урожая, (подробнее: Как влияют внешние факторы на процесс фотосинтеза). Кроме того, необходимы сорта с повышенной интенсивностью фотосинтеза, приспособленные к разным климатическим условиям.
В связи с тем что за счет фотосинтеза используется всего около 1% солнечной радиации, наука занимается проблемой усиления эффективности фотосинтеза и одновременно повышения генетического потенциала продуктивности. До настоящего времени увеличить эффективность фотосинтеза с единицы листовой поверхности с помощью селекции еще не удалось, однако высокой продуктивности добиваются за счет увеличения посевной площади. У некоторых дикорастущих видов из семейства злаковых эффективность фотосинтеза даже выше, чем у культурных видов, что объясняют утолщенным слоем мезофилла листьев, в котором размещены хлоропласты.
Можно предположить, что происходило бы в природе, если бы удалось индуцировать мутации, изменяющие морфологический и биохимический состав хлоропластов, которые получили бы более высокую эффективность фотосинтеза. Между тем такие мутанты скрываются в бесконечно большой по численности популяции соматических клеток. Усовершенствование метода культуры изолированной клетки позволяет обнаружить мутанты с повышенной эффективностью фотосинтеза, что могло бы иметь большое значение для селекции растений.
Дальнейшая возможность заключается в создании растений С3 с системой фотосинтеза, присущей растениям С4. Это также весьма значительное изменение генетической структуры, но обнадеживающих результатов в данном направлении еще не получено. Наряду с открытием и индуцированием генетических изменений необходимо использовать косвенные методы увеличения продуктивности фотосинтеза.
Удлинение вегетационного периода может привести к росту продуктивности суммарной биомассы, а удлинение периода налива зерна или развития плодов - к повышению продуктивности зерна или плодов. Продолжительность вегетационного периода не всегда положительно коррелирует с урожайностью, но удлинение периода налива зерна чаще всего приводит к ее повышению.
Благодаря работе огромного числа генетиков и селекционеров в различных странах мира мы сегодня располагаем коллекциями генотипов, которые включают различные гены, контролирующие продолжительность яровизации, длительность фотопериода, продолжительность периода от цветения до созревания и т.д. Путем благоприятного подбора родительских пар и применения соответствующих методов селекции можно было бы добиться такой рекомбинации генов, которая способствовала бы более продолжительному периоду от цветения до созревания или более раннему началу цветения при неизменном времени созревания или же обеспечила бы трансгрессивное расщепление в обоих направлениях. Все это, разумеется, должно отвечать конкретным агроэкологическим условиям, особенно таким, как высокая температура, засуха или болезни, которые лимитируют формирование зерна.
Одну из возможностей увеличения продуктивности фотосинтеза открывает изменение структуры листьев и стеблей. У культур, выращиваемых в плотном посеве, прямостоячее, или эректофильное, положение листьев дает преимущество по сравнению с плоскостным, или горизонтальным, расположением, поскольку способствует более глубокому прониканию света в глубь посева и к тому же повышает участие и увеличивает продолжительность сроков функционирования листовой поверхности в состоянии фотосинтетической активности. Как уже подчеркивалось ранее, положение листьев под углом около 45° у злаковых трав наряду с их эректофильным положением может быть эффективным в отношении фотосинтеза (рис. 15.7).
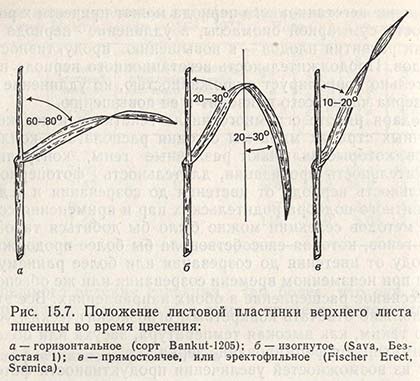
Сорта риса с эректофильными листьями созданы на Филиппинах, гибриды кукурузы с эректофильным положением листьев - в США. В Югославии на основе линии В73, имеющей эректофильные листья (д-р Рассел), выведены Вс-6661, ZP-704, NS SC-606 и другие сорта; они относятся к самым урожайным в своей группе, а это указывает на то, что данный признак способствовал повышению потенциала продуктивности.
Для генотипов пшеницы с эректофильными листьями после колошения характерно сильное сужение верхушки листовых пластинок, однако трудно сказать, в какой мере это влияет на урожайность.
В связи с положением листьев необходимо проанализировать наиболее благоприятное расположение растений в пространстве (густота и способ посева), чтобы получить оптимальный индекс листовой поверхности и выявить формы с более продолжительным периодом функционирования листовой поверхности. В этой главе уже отмечалось, что данный фактор оказывает наибольшее прямое влияние на урожай по сравнению со всеми параметрами зеленой поверхности листьев и может послужить критерием для отбора на урожайность.
Дальнейший путь повышения продуктивности фотосинтеза лежит в создании генетической устойчивости к патогенным организмам. Без этого нельзя максимально использовать генетический потенциал продуктивности существующих сортов и, в частности, предпринимать важные шаги для дальнейшего движения вперед. Генетическая устойчивость к болезням и вредителям имеет огромное значение, и ей была посвящена вся предыдущая глава.
Читайте также: