
Появление цветка у покрытосеменных
Обновлено: 15.09.2024
Покрытосеменные растения — это отдел высших растений, имеющих цветок как орган полового размножения, отличающихся замкнутым вместилищем семяпочки.
Считается, что свою историю на планете покрытосеменные начали 220 млн лет назад — с момента, когда они отделились от гнетовых и образовали отдельное семейство. Примерно 100 млн лет назад отдел уже являлся одним из самых многочисленных и широко распространенных.
Самыми древними из покрытосеменных являются представители группы кувшинковых или нимфейных.
Войти в число доминирующих форм растительной жизни покрытосеменным дала возможность их способность к адаптации. Приспосабливаясь к самым разным условиям существования, они эволюционировали в разнообразные формы: наземные и водные травы, деревья и кустарники, прямостоящие, ползучие и карабкающиеся лианы.
В биологии у этого отдела есть еще одно название — цветковые растения.
Отличительные признаки покрытосеменных
К основным признакам покрытосеменных относятся:
- наличие ограниченных в росте, видоизмененных, приспособленных к размножению спороносных побегов — цветков;
- расположение семязачатков в полости завязи пестика;
- защищенность семян путем погружения в плод;
- способность давать большое потомство, обеспечивать большое количество мутаций;
- двойное оплодотворение;
- многообразие жизненных форм;
- высокая организация проводящей системы.
Также в список характерных особенностей покрытосеменных входит сложный биохимический состав. Все виды цветковых растений вырабатывают целый набор специфических веществ: флавоноиды, эфирные масла, гликозиды, алкалоиды.
Процесс опыления и оплодотворение у цветковых растений
Сначала происходит развитие пыльцы: в пыльниках тычинок производится деление клеток, которые в последующем созревают в пыльцу. Каждое пыльцевое зерно покрыто двухслойной оболочкой. На верхней части этой оболочки имеются особые выросты, с помощью которых зерна удерживаются на рыльце пестика. А внутри размещаются генеративная и вегетативные клетки — это мужской гаметофит.
Затем наступает период развития семязачатков. Они растут на внутренней стороне стенок завязи. Внутри семязачатка формируется зародышевый мешок. Это женский гаметофит. В нем есть яйцеклетка и центральная клетка.
Когда пыльцевое зерно прикрепляется к рыльцу пестика, оно начинается набухать и прорастать, формируя пыльцевую трубку. В ней генеративная клетка делится на два спермия. Затем один спермий сливается с яйцеклеткой, производится оплодотворение и возникновение зиготы. Второй спермий объединяется с центральной клеткой.
Поскольку оплодотворение в данном случае подразумевает слияние двух половых клеток с возникновением зиготы, его называют двойным. Двойное оплодотворение — особая характеристика отдела покрытосеменных.
Сравнение классов однодольные и двудольные
Классификация цветковых предполагает деление на два класса:
- Однодольные — имеющие одну семядолю.
- Двудольные — имеющие две и более семядолей.
В таблице ниже приведены основные отличия в структуре, строении, численности семейств, родов и видов.
Значение покрытосеменных в природе и жизни человека
Значение цветковых в природе переоценить довольно сложно. Благодаря своему разнообразию и многочисленности, они вносят огромный вклад в газообмен экосистемы — вырабатывают большое количество кислорода.
Многие многолетние виды служат домом для животных. Травянистые и кустарниковые выполняют еще одну функцию — являются первичным звеном пищевой цепи.
Представить жизни человека без цветковых тоже достаточно сложно. Они:
- являются важными сельскохозяйственными культурами — например, пшеница, кукуруза, овес, рис, картофель, томаты;
- вносят разнообразие в общий рацион — ягодные и плодовые культуры;
- позволяют получить ткани, выступают сырьем для них.
В ходе технического прогресса человек научился применять многие типы покрытосеменных для производства лекарств. Древесину используют в качестве строительного материала. А декоративные виды — для ландшафтного дизайна.
Механизмы реализации основных жизненных функций
Все покрытосеменные имеют вегетативные и репродуктивные органы. К вегетативным относят:
Эти органы выполняют всю работу по бесполому размножению.
К генеративным — реализующим схему полового размножения относят:
Семена формируются по окончании периода цветения. Они надежно защищены околоплодником. Поэтому хорошо сохраняются и легко распространяются.
Семена цветковых очень сильно различаются по размеру, цвету и форме. Например, семена пшеницы имеют размер 27–30 мкм, вес 35 мг. А семя сейшельской пальмы весит 10 кг, вытягивается в длину до 50 см.
Важнейшую роль в реализации жизненных функций играет цветок. Он привлекает агентов опыления — насекомых — ярким цветом и ароматом пыльцы. Ботаники считают, что цветок является ответом на вопрос, почему именно покрытосеменные стали доминирующей формой растительной жизни и смогли приспособиться к самым разным климатическим и природным условиям.

Цветковые, или покрытосеменные – сравнительно молодая группа высших растений, появившаяся в меловом периоде, стремительно размножившаяся в середине – конце мела и с тех пор занимающая господствующее положение на суше. Проблеме происхождения цветковых посвящено огромное количество публикаций, однако в этом вопросе по-прежнему много неясного. Общепризнано, что цветковые произошли от голосеменных, но от кого именно и каким образом?
Д.Д.Соколов и А.К.Тимонин с кафедры высших растений биофака МГУ подробно рассмотрели основные теории происхождения покрытосеменных в обзорной статье, опубликованной во втором номере Журнала общей биологии за 2007 г.
Авторы обращают внимание на парадоксальный факт: цветы покрытосеменных до такой степени разнообразны, что не существует ни одного признака или структурной особенности, которая была бы присуща всем цветкам без исключения. Поэтому и сформулировать, что такое цветок, весьма трудно. Тем не менее ясно, что именно появление цветка было ключевым событием в происхождении покрытосеменных.
Впрочем, если не рассматривать мужские цветки, у остальных все-таки есть общая черта, отличающая цветок покрытосеменных от фруктификаций голосеменных. У цветковых плодолистики свернуты и образуют, во-первых, рыльце, улавливающее пыльцу, во вторых – замкнутую полость (завязь), в которой развиваются семязачатки. У некоторых голосеменных есть нечто похожее – замкнутые капсулы с семязачатками, однако у них нет завязи (пыльцу улавливают сами семязачатки при помощи длинных отростков, высовывающихся из узкого отверстия капсулы). К тому же исходный листочек, несущий семязачатки ( мегаспорофилл , у цветковых ему соответствует плодолистик) свернут у голосеменных нижней (абаксиальной) стороной внутрь, а у цветковых – наоборот, верхней (адаксиальной) стороной внутрь. Соответственно и семязачатки сидят на разных сторонах листочка.
В начале XX века была популярна теория происхождения цветковых от беннеттитов – вымершей группы голосеменных, у которых имелись обоеполые фруктификации (стробилы) – в отличие от других голосеменных, имеющих однополые фруктификации. Из стробилов беннеттитов, действительно напоминающих цветы, ботаники и пытались вывести типичный для покрытосеменных обоеполый цветок. Но тут возникла проблема: у беннеттитов не обнаружилось никаких структур, из которых можно было бы вывести плодолистики. У них вообще нет мегаспорофиллов, а семязачатки сидят прямо на оси стробила. Проблема казалась неразрешимой.

Ничего невероятного в таком сценарии нет, исходя из того, что известно об общих принципах генетической регуляции развития. Гипотеза Мейена исключительно изящно и просто объясняет противоречие, казавшееся неразрешимым. Ее нельзя проверить классическими методами палеоботаники и морфологии (поскольку никаких переходных форм гипотеза Мейена не предполагает). Однако ее можно проверить методами сравнительно-генетического анализа.
Эволюционные реконструкции, основанные на морфологии, подтверждали родство покрытосеменных и беннеттитовых (а также родственных беннеттитам гнетовых, которые дожили до наших дней). Однако реконструкции, основанные на молекулярных данных, показали иное. Они показали, что все дожившие до современности голосеменные представляют собой монофилетичную группу (т.е. группу, происходящую от одного общего предка и включающую всех без исключения потомков этого предка). Это значит, что последний общий предок всех современных голосеменных, в том числе гнетовых (а стало быть и беннеттитов тоже) не был предком покрытосеменных. Таким образом, предка цветковых нужно искать среди других вымерших голосеменных, более древних, точнее, обособившихся раньше, чем беннеттитовые. Фролик и Паркер предложили свою кандидатуру – користоспермовые семенные папоротники (Corystospermataceae). У них, правда, микроспорангии (пыльники) располагались на нижней, а не на верхней стороне микроспорофиллов.
Авторы предполагают, что причина всех этих противоречий, вероятно, кроется в недостаточной надежности эволюционных реконструкций, основанных на молекулярных данных. По их мнению, голосеменные растения плохо подходят для молекулярно-филогенетического анализа, поскольку большинство семейств голосеменных давно вымерло. Мы, таким образом, имеем недостаточно репрезентативную выборку таксонов, что, как хорошо известно, может приводить к ошибочным молекулярным филогениям. В конечном счете, по мнению авторов, на сегодняшний день нет оснований для безоговорочного отрицания идеи Мейена о происхождении цветковых от беннеттитовых. Эту идею вполне можно сохранить в качестве рабочей гипотезы.
Представляют ли цветковые растения одну большую естественную группу, происходящую от одного общего корня, или они состоят из нескольких или даже многих независимых линий развития, имеющих происхождение от разных предков? Исключительно большое разнообразие внешней формы и внутреннего строения цветковых растений (сравнить только ряску с дубом!) иногда дает повод к предположению о независимом происхождении разных групп цветковых от разных голосеменных предков, а иногда даже от разных отделов высших растений. Однако эта идея так называемого полифилетического происхождения цветковых растений противоречит современным данным об их строении, развитии и систематике. Общность происхождения всех известных нам цветковых растений убедительно доказывается прежде всего множеством общих внешнеморфологических, анатомических и эмбриологических, признаков между представителями самых различных семейств и порядков, в том числе и таких признаков, которые не связаны между собой в онтогенезе и в процессе эволюции. Общность происхождения доказывается, в частности, общностью для цветковых растений тройного слияния с образованием характерного только для них триплоидного эндосперма. За естественность отдела цветковых растений, за его происхождение от общего предка говорят также данные современной систематики. Даже самые своеобразные и в систематическом отношении кажущиеся совершенно обособленными цветковые растения, как знаменитая австралийская казуарина и внешне похожий скорее на гриб паразитный род баланофора, связаны с теми или иными группами через промежуточные звенья. Все те группы, которые кажутся стоящими совершенно изолированно и внушают мысль, особенно людям, недостаточно знакомым с миром цветковых растений в целом, о независимом, полифилетическом происхождении, при ближайшем исследовании и более широком сравнении с другими группами рано или поздно находят себе естественное место в системе отдела цветковых. Поэтому среди систематиков, занимающихся разработкой системы всего отдела цветковых растений, в настоящее время уже нет сторонников их полифилетического происхождения. Интересно, что точка зрения систематиков подтверждается новейшими данными молекулярной биологии, в частности изучением последовательности аминокислот в дыхательном ферменте цитохроме С.
Но не могли ли цветковые растения произойти от наиболее примитивных голосеменных — семенных папоротников, у которых, как известно, стробилов еще небыло? В настоящее время ряд ботаников, занимающихся вопросом о происхождении цветковых растений, приходят к выводу, что неизвестные пока еще нам непосредственные предки цветковых растений были, вероятно, тесно связаны с семенными папоротниками и, возможно, представляли собой одну из ветвей этой примитивной группы. Одним из веских доводов в пользу вероятности происхождения цветковых растений от семенных папоротников является так называемый внешний интегумент семязачатков. Как читатель знает уже об этом из раздела о строении семязачатка, по мнению ряда ботаников, внешний интегумент произошел скорее всего из так называемой кунулы:, которая была характерна для некоторых более подвинутых групп семенных папоротников, таких, как медуллозовые, користоспермовые и кейтониевые.
Таким образом, вопрос о предках цветковых растений все еще не вышел за рамки более или менее правдоподобных гипотез. Осадочные горные породы упорно хранят молчание, и пока нет намеков на возможность скорого разрешения этой проблемы. Более того, если даже в один прекрасный день будут фактически найдены ископаемые остатки предков цветковых растений, есть очень большая опасность, что мы далеко не сразу сможем установить их родственную связь с цветковыми. Очень важно, чтобы были открыты не только остатки вегетативных частей и не только пыльцевые зерна или изолированные от растения семена или даже плоды, но и цветок со всеми его основными органами. Но в геологических отложениях остатки цветков встречаются значительно реже, чем другие более стойкие к разрушению части растеиия. Тем не менее такие находки иногда делаются, и не исключено, что когда-нибудь будет найдено некое архаическое цветкоподобное образование, которое ботаники сочтут возможным истолковать как архетип цветка.
В раннем мелу цветковые растения играли лишь ничтожную роль в растительном покрове Земли и встречались довольно редко. Однако к концу альба, т. е. в середине мелового периода (приблизительно 100 млн. лет назад), происходит одно из наиболее глубоких и резких изменений растительного мира суши и цветковые растения за сравнительно короткий промежуток геологического времени, исчисляемого несколькими миллионами лет, широко распространяются по всему земному шару и быстро достигают Арктики и Антарктики. Одним из основных условий их быстрого распространения была, вероятно, их высокая эволюционная пластичность, что выражается в необычайном разнообразии многочисленных приспособлений к самым различным экологическим условиям. Все другие представители нижнемеловой флоры отличались, по-видимому, значительно меньшей приспособляемостью к быстро изменяющимся экологическим условиям. Чрезвычайно большое значение для быстрой эволюции, и широкого массового расселения цветковых растений имели насекомые как опылители и птицы как агенты распространения семян, а затем и плодов, В результате быстрой адаптивной радиации цветковые растения оказались способными к образованию большого разнообразия сообществ, входящих в состав самых различных экосистем. В отличие от голосеменных, среди которых неизвестны настоящие травянистые формы, у цветковых, притом в самых различных линиях эволюции, возникло очень большое разнообразие трав, в том числе много эпифитных форм. Цветковые оказались единственной группой растений, способной к образованию сложных многоярусных сообществ, состоящих главных образом, а иногда почти исключительно, из самих цветковых. Возникновение таких сложных многоярусных сообществ не могло не способствовать как более интенсивному и полному использованию среды, так и более успешному завоеванию новых территорий и колонизации новых местообитаний.
1. Несмотря на то что в обширных областях Западной Гондваны (в Африке и особенно в Южной Америке) мезофитная тропическая лесная флора сохранилась достаточно хорошо и в очень большом разнообразии форм, в Юго-Восточной Азии и соседних областях сохранилось наибольшее число примитивных форм, гораздо больше, чем в Америке и особенно в Африке.
2. В тех случаях, когда примитивные группы сохранились как в Юго-Восточной Азии и соседних областях, так и на территории, соответствующей Западной Гондване, в Западной Гондване они представлены в меньшем разнообразии и более подвинутыми представителями. Так, примитивные семейства магнолиевых и винтеровых отсутствуют в Африке (и только очень специализированный монотипный род Takhtajania из винтеровых встречается на Мадагаскаре), а в Америке представлены меньшим числом родов (3 рода из 12 родов магнолиевых и 1 род винтеровых) и видов и менее примитивными таксонами, чем в Юго-Восточной Азии и соседних областях.
3. В Юго-Восточной Азии и соседних областях произрастает не только много примитивных семейств, но и наиболее примитивные представители многих более подвинутых семейств и родов, как двудольных, так и однодольных.
Сравнительное изучение примитивных ныне живущих цветковых растений позволяет также делать некоторые выводы об облике и некоторых морфологических особенностях ранних цветковых растений. Ни одно из ныне живущих цветковых не обладает всеми примитивными признаками, так как все, даже относительно самые примитивные таксоны, специализировались в том или ином направлении. Однако, суммируя все наиболее архаические признаки, рассеянные среди магнолиевых, винтеровых, дегенериевых и других примитивных семейств, можно с определенной степенью вероятности воссоздать некоторые черты ранних цветковых растений. Можно с достаточной уверенностью утверждать, что это были древесные растения (Ч. Джеффри, 1899, 1917), скорее всего небольшие деревья (X. Халлир, 1901, 1912) или, быть может, даже кустарники. Этот вопрос трудно решить сколько-нибудь уверенно, так как среди современных примитивных покрытосеменных встречаются как относительно высокие деревья, так и кустарники (виды тасманнии и эвпоматии). Но если правильна гипотеза, что цветковые произошли в условиях экологического стресса, то это скорее всего должны были быть небольшие кустарники, но не полукустарники, как думает Стеббинс (1974), так как полукустарники — это, несомненно, вторичная жизненная форма. Ксилема была лишена сосудов. Листья были вечнозеленые (как почти у всех голосеменных), очередные, вероятно, более или менее ксероморфные, с перистым жилкованием и парацитными устьицами (с побочными клетками, расположенными по обе стороны от замыкающих клеток, параллельно их длинной оси). Цветки ранних цветковых были, вероятно, в примитивных верхоцветных соцветиях. Они были обоеполые, с умеренно удлиненным цветоложем, на котором в спиральном порядке располагались чашелистики, тычинки и плодолистики. Первоначально цветки были, вероятно, лишены лепестков, которые возникли позднее, главным образом из тычинок. Цветки опылялись насекомыми, скорее всего жуками. Пыльцевые зерна были одиобороздные, с гладкой экзиной и еще без характерного для цветковых, растений столбикового слоя в эктэкзине. Семена были, вероятно, с сильно развитым слоем из живых паренхимных клеток и. распространялись птицами. Плоды представляли собой многолистовку. Если бы этот гипотетический первичный тип цветкового растения сохранился (или был бы найден в ископаемом состоянии), то систематики отнесли бы его скорее всего к порядку магнолиевых.
По всем данным, эволюция цветковых с самого начала происходила путем широкой адаптивной радиации и шла очень быстрыми темпами, что объясняется как экологическими, так и генетическими и цитогенетическими факторами (в частности, большой ролью хромосомных перестроек и полиплоидизации). В результате уже к середине мелового периода цветковые достигли очень большого разнообразия форм и оказались приспособленными к самым различным экологическим условиям, к возрастающему разнообразию экологических ниш. К этому времени уже возникли все основные систематические группы цветковых растений вплоть до многих родов. Начиная с середины мелового периода цветковые занимают доминирующее положенно.
Отдел цветковых, или покрытосеменных, растений — это самая многочисленная и хорошо приспособленная к современным условиям группа представителей флоры. Они довольно разнообразны и включают в себя травы, кустарники и деревья. Их основной отличительной чертой является наличие цветка как органа размноженья, а также замкнутого вместилища у семян, откуда и происходит название отдела.

Происхождение и эволюция
Считается, что предком этой группы растений были водоросли, не сохранившиеся до современности. Затем они эволюционировали в папоротники, которые в свою очередь дали начало как покрытосеменным, так и голосеменным видам Они произошли от общих предков, разделившихся на разные ветви в триасе.

Всего цветковые существуют около 140 миллионов лет — первые представители фауны с похожим строением появились в юрском периоде. Это были довольно примитивные и малочисленные виды. Древнейшими представителями цветковых считаются представители семейства нимфейные или кувшинковые. В середине мелового периода (около 100 миллионов лет назад) покрытосеменные уже были довольно широко распространены в мире. В конце мела они оказались доминирующей формой жизни и покрывали значительную часть суши. Примерно тогда же начали появляться предки многих современных растений — дуба, клёна, магнолии, бука. Одной из главной особенностей эволюции цветковых стала их высокая приспосабливаемость к изменяющимся условиям, что позволило им произрастать на разных географических широтах. Они отличаются огромным многообразием форм, размеров, способов развития и размножения. Среди цветковых можно найти водные и наземные, ползучие, вьющиеся, карабкающиеся и прямостоящие растения, которые растут и процветают во всех уголках Земли, от тропических лесов и долин рек до солёных морей и альпийских лугов. Другой интересной особенность эволюции покрытосеменных является большое разнообразие форм водяных растений. Они не произошли от водорослей напрямую, а эволюционировали от наземного предка в результате смены условий, потому отличаются сложным строением и большим количеством приспособительных механизмов.
Основные признаки
Для того чтобы дать краткую характеристику покрытосеменным растениям, нужно посмотреть на их основные признаки. Их список выглядит так:

- растение состоит из стебля, корневой системы, листьев и цветка;
- семя защищено околоплодником — оболочкой, которая обеспечивает его сохранность при распространении;
- семязачатки и семена хранятся в завязи и плоде;
- женский заросток представляет собой зародышевый мешок с восемью ядрами, а мужской — пыльцевое зерно, состоящее из вегетативных и генеративных клеток;
- двойное оплодотворение — спермии (мужские гаметы) цветка воздействуют одновременно на яйцеклетку и ядро зародышевого мешка;
- для размножения растению необходимо опыление, которое может осуществляться по воздуху или воде, а также с помощью переноса пыльцы птицами и насекомыми;
- завязь оплодотворённого цветка преобразуется в плод, семяпочка — в семя, зигота — в зародыш семени, вторичное ядро — в эндосперм;
- половое размножение с постоянной сменой поколений.
К другим типичным признакам покрытосеменных относятся ситовидные трубки на стебле, широкие пластинчатые листья со сложной системой сетчатых жилок, цветки циклической формы. Но стоит отметить, что ни один из характерных признаков группы не свойственен всем без исключения её членам. Принадлежность вида к цветковым определяется по общему числу признаков, но некоторые из них могут отсутствовать.
Классы покрытосеменных
Всех представителей отдела покрытосеменных делят на однодольные и двудольные растения. Как можно понять из их названия, они отличаются друг от друга по количеству семядолей. Сравнительная характеристика этих классов:
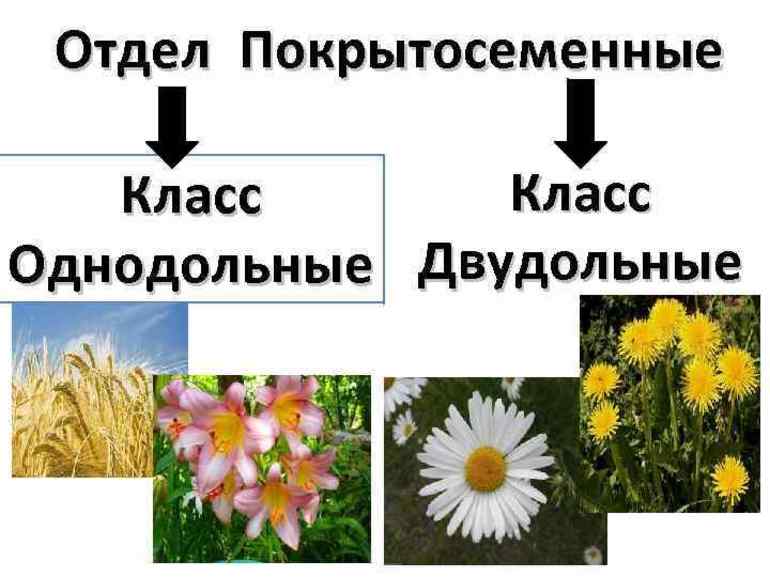
- для однодольных характерна корневая система мочковатой формы, состоящая в основном из придаточных отростков, а для двудольных — мощный главный корень в форме стержня;
- листья растений первого класса имеют простую структуру и состоят из обычной цельной пластинки, в то время как у второго они могут иметь более сложное строение;
- жилкование у однодольных параллельное или дуговое, а у двудольных — сетчатое;
- для растений с одной семядолей характерно количества частей цветка, кратное трём, а с двумя — четырём или пяти;
- у однодольных покрытосеменных нет камбия — образовательной ткани, обеспечивающей рост стебля и корней в толщину;
- проводящие пучки у двудольных расположены в виде кольца, а у родственного класса — беспорядочно и хаотично.
В класс двудольных входят высокоразвитые деревья и кустарники, в том числе многочисленные плодовые и декоративные культуры. Их общее количество достигает 180—190 тысяч.
Семейства однодольных
Всего в современной классификации насчитывается более 500 семейств покрытосеменных, из которых 12 — двудольные. Они отличаются строением стебля и листьев, формулой цветка, длительностью жизненного цикла, местами, где они предпочитают расти и многими другими особенностями. Глядя на фото и названия цветковых растений разных видов, трудно не поразиться их разнообразию — в один отдел входят самые разные формы жизни, от водяной ряски до огромного дуба.

- стебель-соломина;
- небольшие корни с развитыми придатками;
- сворачивающиеся при жаре листки;
- простые цилиндроподобные цветки, собранные в злаки;
- сухие невскрывающиеся плоды;
- опыление ветром.

- подземные луковицы или клубнелуковицы, которые формируются из побегов и могут защитить почки от неблагоприятного воздействия внешней среды во время жары заморозков;
- прямой или вьющийся надземный стебель, который называется цветочной стрелкой;
- узкие мясистые листья;
- широкое разнообразие соцветий;
- цветки обоеполые;
- плод-коробочка или ягода;
- опыление насекомыми, в тропических широтах также птицами, редко — ветром.
К примерам цветковых растений из семейства лилейных можно отнести самые разные виды, от ландышей до лука. Бо́льшую часть из них составляют многолетние травы, реже встречаются деревья и кустарники.
Двудольные растения
Среди группы двудольных растений выделяется класс розоцветных. Эти растения насчитывают 300 видов в 100 родах. Распространены в субтропиках и местах с умеренным климатом. Отличаются особым строением цветоложа. Опыляются насекомыми, привлекая их нектаром или выделяя очень большое количество пыльцы, чтобы размножаться. Все представители класса имеют очень похожие цветки. Но их плоды отличаются широким разнообразием — среди них можно найти ягоды, орешки, листовки, яблоки, коробочки. Многие из них приспособлены к распространению животными.

Семейство бобовых получило своё название за плоды в виде бобов или стручков. Оно состоит из 24,5 тысяч растений, объединённых в 946 родов. К особенностям, которые могут указать на принадлежность к этому классу, относятся стоячие вьющиеся стебли, сложные по строению листья с прилистником и цветы из пяти лепестков. Цветы обоеполые. Опыляются насекомыми. К семейству паслёновых относятся травы, кусты и маленькие деревья. К особенностям, которые могут показать принадлежность вида к ним, относится расположение листьев. В вегетативной части они растут попеременно, а в цветоносной — попарно. Из-за этого стебли у представителей флоры кажутся разветвлёнными на две или три части. Другие особенности семейства — соцветия в форме завитков, небольшое количество тычинок равной длины, похожие на почки семена, плоды в виде ягод или коробочек.

Крестоцветные растения разделены на 4 тысячи видов, которые включают 400 разновидностей. В семейство входит много двулетних монокарпических растений с особой схемой цветения — они размножаются только один раз на последнем году жизни. К особенностям также относится стержневая корневая система (реже встречаются корнеплоды), простые листья без прилистника, толстый стебель, 6 тычинок, плод-стручок и кистеобразные соцветия. Ещё одним интересным семейством двудольных растений являются астровые, или сложноцветные. Это одна из самых больших групп, которая насчитывает почти 40 тысяч видов в 1911 родах. Их можно найти в любом уголке земного шара, от тундры до тропиков. Основной особенностью семейства является соцветие-корзинка, которое образуется из множества очень мелких цветков. Кроме того, для них характерен развитый корень-стержень и плод-семянка, не содержащий белка.
Хозяйственное значение
Трудно переоценить то, сколько пользы человеку приносят покрытосеменные растения. Их используют в следующих сферах жизни:

- пищевая промышленность, изготовление кормов для животных;
- медицина — как лекарственные растения в составе различных препаратов;
- дизайн интерьера и ландшафта — как декорации;
- ремесло — для изготовления духов, косметических средств, красителей, масел и прочего;
- в культуре как подарки и сувениры;
- в науке для исследований генетики.
Экологическое положение
Сегодня покрытосемянные являются самыми многочисленным растениями, господствующими по всему земному шару. Чтобы понять, почему они смогли занять эту нишу, достаточно посмотреть на их эволюционный путь — эти представители флоры развивались в мезозойскую и кайнозойскую эру, когда условия были очень близки к современным.

Эти растения получили центральную нишу в экосистемах и в какой-то мере повлияли на эволюцию почти всех животных и птиц, для которых они были источником пищи и основой среды обитания. Между тем большую роль в этом деле сыграла и деятельность человека — в частности, окультуривание растений и выведение новых сортов, приспособленных под разные условия.
Несмотря на это, отдельные виды покрытосеменных находятся под угрозой исчезновения и занесены в Красную книгу. Причиной этого является в основном деятельность людей — вырубка лесов, пожары, осушение и затопление зон для ведения сельского хозяйства, накопление газа в атмосфере.
Покрытосеменные растения — наиболее распространённая и разнообразная группа представителей флоры. Они играют очень важную роль как в жизни человека, так и в развитии экосистем. Разные классы, подклассы, семейства и виды цветковых могут очень сильно отличаться друг от друга и не иметь почти никаких общих внешних черт и особенностей жизнедеятельности, кроме защищённого околоплодником семени.
Читайте также: