
Черная субстанция отделяет базальную часть ножек мозга от
Обновлено: 08.07.2024
Базальные ганглии обычно изоденсивные или изотенсивные по отношению к коре головного мозга. Бледный шар содержит больше миелина по сравнению со скорлупой, поэтому бледное ядро визуализируется более гипоинтенсивным на T2WI, GRE и SWI изображениях. При старении в бледном шаре откладывается кальций, что приводит к снижению сигнала, а при кальцификации более, чем на 40% сигнал теряется на всех последовательностях. При старении также в скорлупе откладывается железо, что приводит к повышению сигнала на Т2. Данное явление часто выявляется у пациентов в возрасте от 70 до 80 лет.
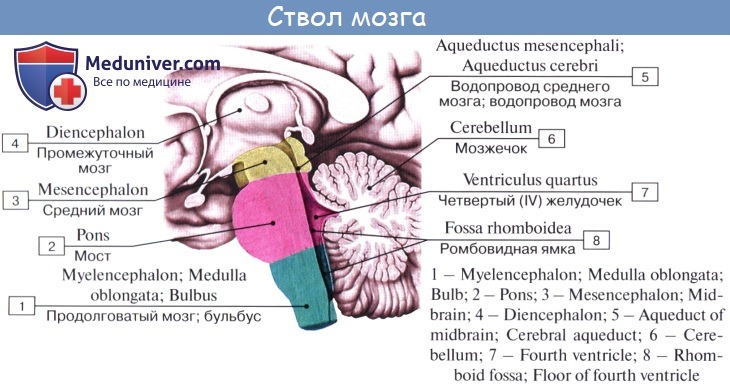
Средний мозг, mesencephalon, развивается в процессе филогенеза под преимущественным влиянием зрительного рецептора, поэтому важнейшие его образования имеют отношение к иннервации глаза. Здесь же образовались центры слуха, которые вместе с центрами зрения в дальнейшем разрослись в виде четырех холмиков крыши среднего мозга.
С появлением у высших животных и человека коркового конца слухового и зрительного анализаторов в коре переднего мозга слуховые и зрительные центры среднего мозга сами попали в подчиненное положение и стали промежуточными, подкорковыми. С развитием у высших млекопитающих и человека переднего мозга через средний мозг стали проходить проводящие пути, связывающие кору конечного мозга со спинным (ножки мозга).
В результате в среднем мозге человека имеются:
1) подкорковые центры зрения и ядра нервов, иннервирующих мышцы глаза;
2) подкорковые слуховые центры;
3) все восходящие и нисходящие проводящие пути, связывающие кору головного мозга со спинным и идущие транзитно через средний мозг;
4) пучки белого вещества, связывающие средний мозг с другими отделами центральной нервной системы.
Соответственно этому средний мозг, являющийся у человека наименьшим и наиболее просто устроенным отделом головного мозга, имеет две основные части: крышу, где располагаются подкорковые центры слуха и зрения, и ножки мозга, где преимущественно проходят проводящие пути.
Дорсальная часть, крыша среднего мозга, tectum mesencephali.
Она скрыта под задним концом мозолистого тела и подразделяется посредством двух идущих крест-накрест канавок — продольной и поперечной — на четыре холмика, располагающихся попарно.
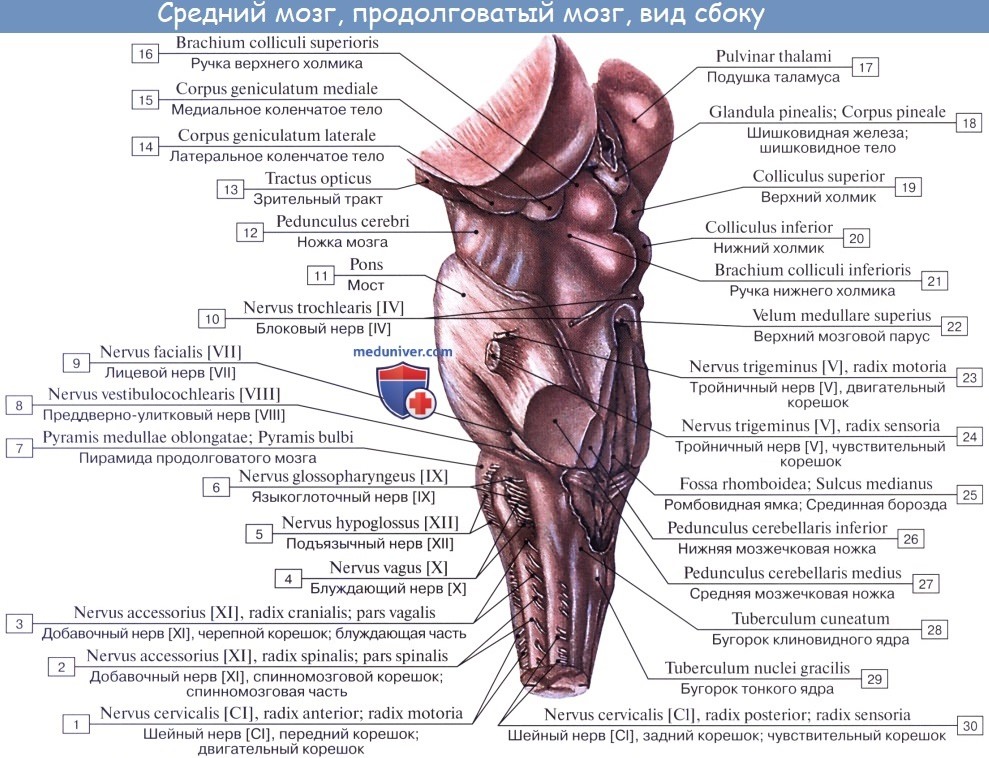
Верхние два холмика, colliculi superiores, являются подкорковыми центрами зрения, оба нижних, colliculi inferiores,— подкорковыми центрами слуха. В плоской канавке между верхними бугорками лежит шишковидное тело. Каждый холмик переходит в так называемую ручку холмика, brachium colliculi, направляющуюся латерально, кпереди и кверху, к промежуточному мозгу. Ручка верхнего холмика, brachium colliculi superioris, идет под подушкой, pulvinar, таламуса к латеральному коленчатому телу, corpus geniculatum laterale.
Ручка нижнего холмика, brachium colliculi inferioris, проходя вдоль верхнего края trigonum lemnisci до sulcus lateralis mesencephali, исчезает под медиальным коленчатым телом, corpus geniculatum mediale. Названные коленчатые тела относятся уже к промежуточному мозгу.
Вентральная часть, ножки мозга, pedunculi cerebri, содержит все проводящие пути к переднему мозгу.
Ножки мозга имеют вид двух толстых полуцилиндрических белых тяжей, которые расходятся от края моста под углом и погружаются в толщу полушарий большого мозга.
Полость среднего мозга, являющаяся остатком первичной полости среднего мозгового пузыря, имеет вид узкого канала и называется водопроводом мозга, aqueductus cerebri. Он представляет узкий, выстланный эпендимой канал 1,5 — 2,0 см длиной, соединяющий IV желудочек с III. Дорсально водопровод ограничивается крышей среднего мозга, вентрально — покрышкой ножек мозга.
На поперечном разрезе среднего мозга различают три основные части:
1) пластинку крыши, lamina tecti;
2) покрышку, tegmentum, представляющую верхний отдел pedunculi cerebri;
3) вентральный отдел pedunculi cerebri, или основание ножки мозга, basis pedunculi cerebralis.
Соответственно развитию среднего мозга под влиянием зрительного рецептора в нем заложены различные ядра, имеющие отношение к иннервации глаза.
У низших позвоночных верхнее двухолмие служит главным местом окончания зрительного нерва и является главным зрительным центром. У млекопитающих и у человека с переносом зрительных центров в передний мозг остающаяся связь зрительного нерва с верхним холмиком имеет значение только для рефлексов. В ядре нижнего холмика, а также в медиальном коленчатом теле оканчиваются волокна слуховой петли (lemniscus lateralis). Крыша среднего мозга имеет двустороннюю связь со спинным мозгом — tractus spinotectalis и tractus tectobulbaris et tectospinalis. Последние после перекреста в покрышке идут к мышечным ядрам в продолговатом и спинном мозге. Это так называемый зрительно-звуковой рефлекторный путь, о котором говорилось при описании спинного мозга. Таким образом, пластинку крыши среднего мозга можно рассматривать как рефлекторный центр для различного рода движений, возникающих главным образом под влиянием зрительных и слуховых раздражений.
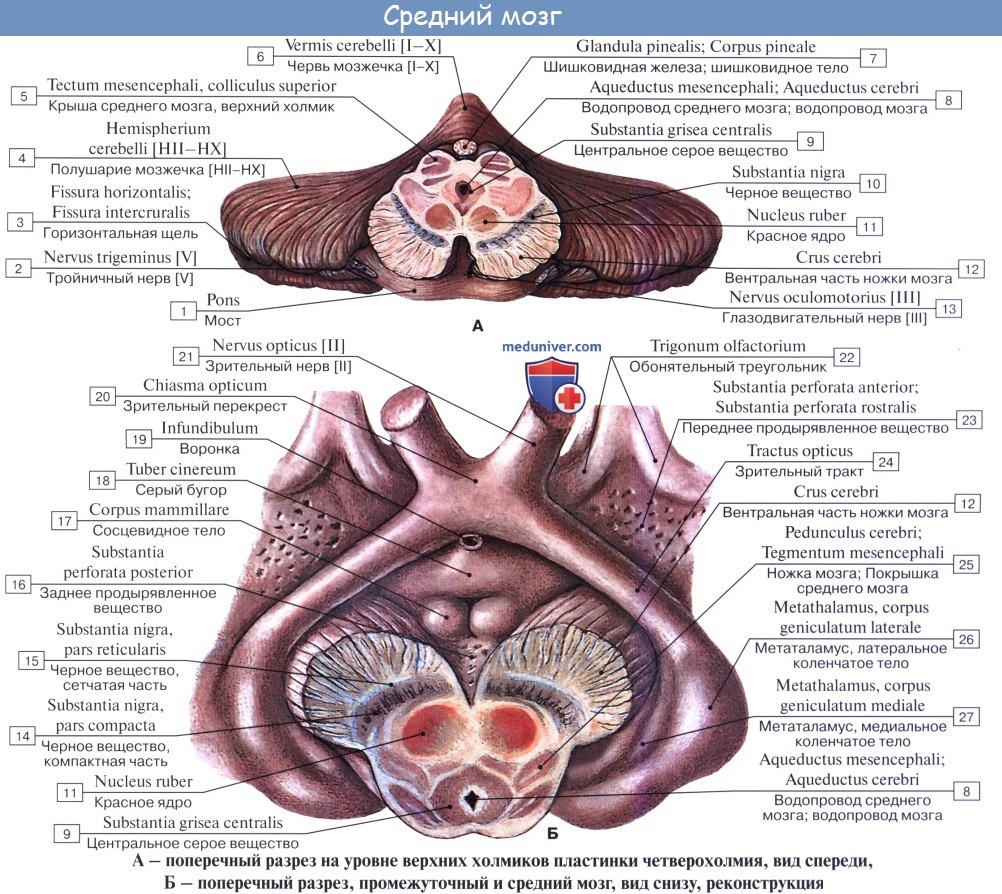
Водопровод мозга окружен центральным серым веществом, имеющим по своей функции отношение к вегетативной системе. В нем, под вентральной стенкой водопровода, в покрышке ножки мозга заложены ядра двух двигательных черепных нервов — n. oculomotorius (III пара) на уровне верхнего двухолмия и n. trochlearis (IV пара) на уровне нижнего двухолмия. Ядро глазодвигательного нерва состоит из нескольких отделов соответственно иннервации нескольких мышц глазного яблока.
Медиально и кзади от него помещается еще небольшое, тоже парное, вегетативное добавочное ядро, nucleus accessories, и непарное срединное ядро. Добавочное ядро и непарное срединное ядро иннервируют непроизвольные мышцы глаза, m. ciliaris и m. sphincter pupillae. Эта часть глазодвигательного нерва относится к парасимпатической системе. Выше (ростральнее) ядра глазодвигательного нерва в покрышке ножки мозга располагается ядро медиального продольного пучка.
Латерально от водопровода мозга находится ядро среднемозгового тракта тройничного нерва, nucleus mesencephalicus n. trigemini.
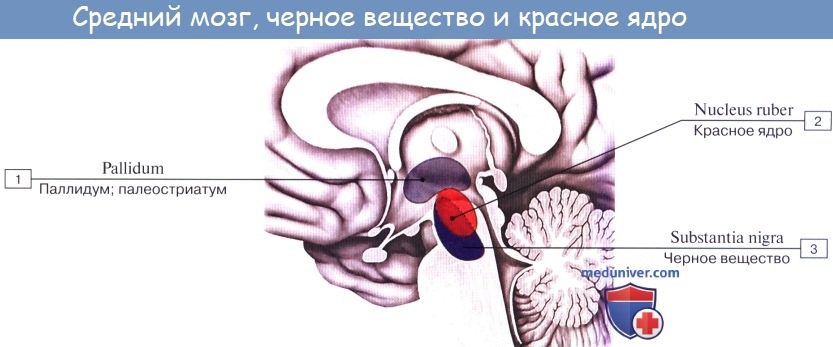
Ножки мозга делятся, как уже отмечалось, на вентральную часть, или основание ножки мозга, basis pedunculi cerebralis, и покрышку, tegmentum. Границей между ними служит черное вещество, substantia nigra, обязанное своим цветом содержащемуся в составляющих его нервных клетках черному пигменту — меланину.
Покрышка среднего мозга, tegmentum mesencephali, — часть среднего мозга, расположенная между его крышей и черным веществом (substantia nigra) ножек мозга.
От нее отходит tractus tegmentalis centralis — центральный покрышечный путь — проекционный нисходящий нервный путь, расположенный в центральной части покрышки среднего мозга. Он содержит волокна, идущие от таламуса, бледного шара, красного ядра и ретикулярной формации среднего мозга к ретикулярной формации и оливе продолговатого мозга; относится к экстрапирамидной системе.
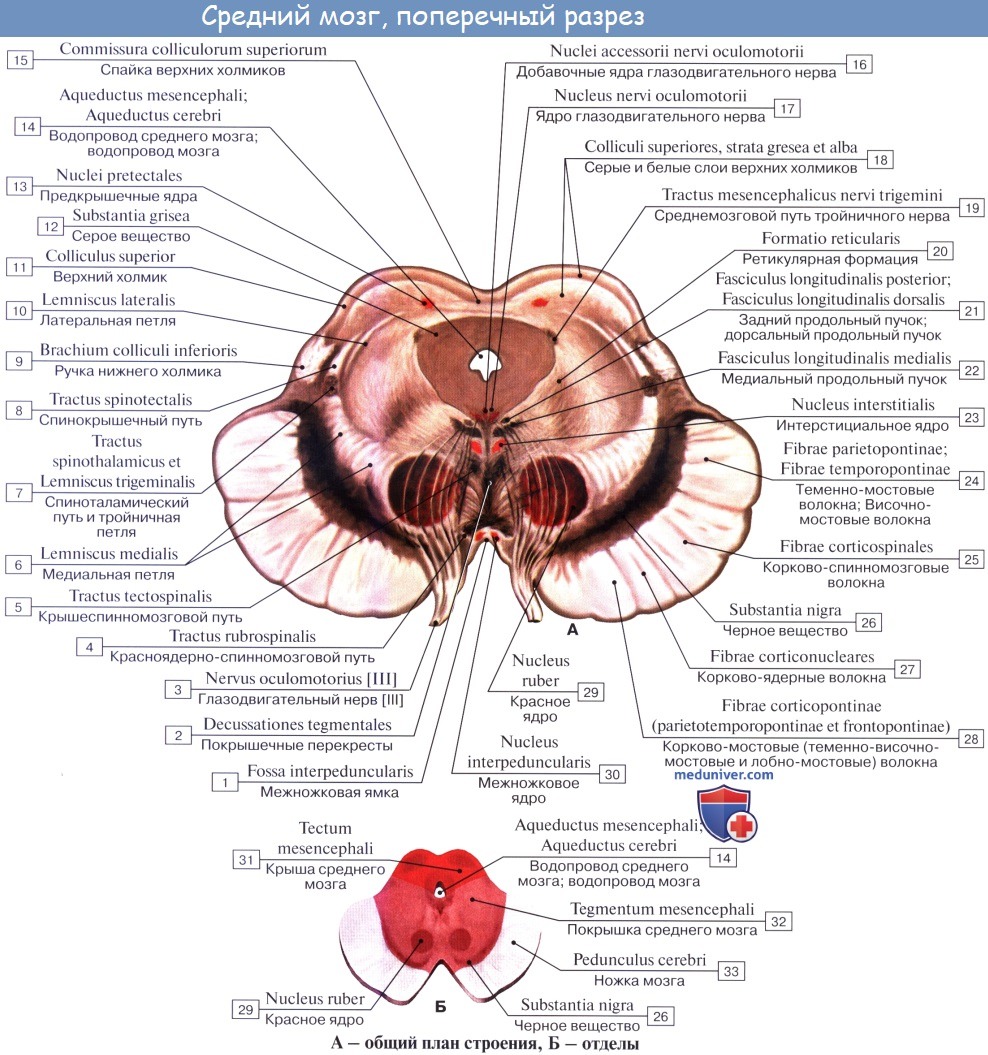
Substantia nigra простирается на всем протяжении ножки мозга от моста до промежуточного мозга; по своей функции относится к экстрапирамидной системе.
Расположенное вентрально от substantia nigra основание ножки мозга содержит продольные нервные волокна, спускающиеся от коры полушария большого мозга ко всем нижележащим отделам центральной нервной системы (tractus corticopontmus, corticonuclearis, corticospinalis и lдр.).
Tegmentum, находящаяся дорсально от substantia nigra, содержит преимущественно восходящие волокна, в том числе медиальную и латеральную петли. В составе этих петель восходят к большому мозгу все чувствительные пути, за исключением зрительного и обонятельного.
Среди ядер серого вещества самое значительное — красное ядро, nucleus ruber. Это удлиненное колбасовидное образование простирается в покрышке ножки мозга от гипоталамуса промежуточного мозга до нижнего двухолмия, где от него начинается важный нисходящий тракт, tractus rubrospinal, соединяющий красное ядро с передними рогами спинного мозга. Пучок этот после выхода из красного ядра перекрещивается с аналогичным пучком противоположной стороны в вентральной части срединного шва — вентральный перекрест покрышки.
Nucleus ruber является весьма важным координационным центром экстрапирамидной системы, связанным с остальными ее частями. К нему проходят волокна от мозжечка в составе верхних ножек последнего после их перекреста под крышей среднего мозга, вентрально от aqueductus cerebri, а также от pallidum — самого нижнего и самого древнего из подкорковых узлов головного мозга, входящих в состав экстрапирамидной системы. Благодаря этим связям мозжечок и экстрапирамидная система через посредство красного ядра и отходящего от него tractus rubrospinal оказывают влияние на всю скелетную мускулатуру в смысле регуляции бессознательных автоматических движений.
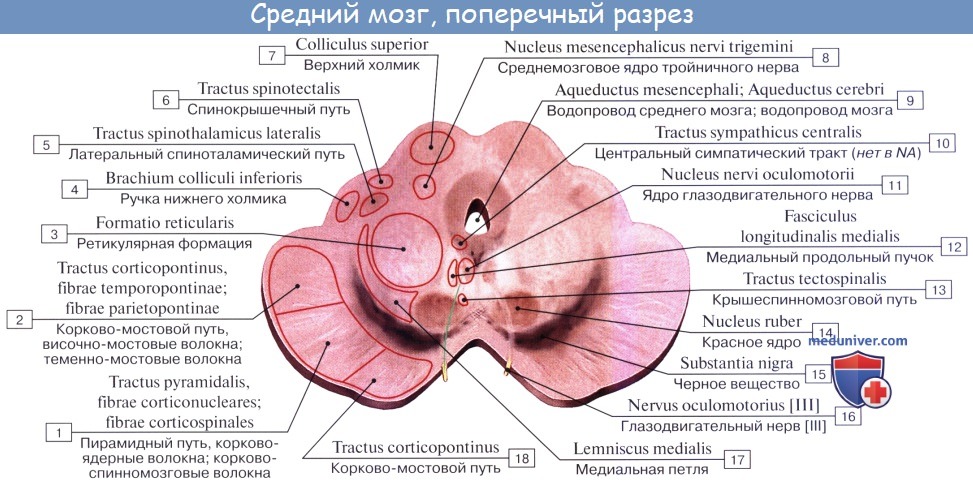
В покрышку среднего мозга продолжаются также ретикулярная формация, formatio reticularis, и fasciculus longitudindlis medialis. Последний берет начало в различных местах. Одна из его частей начинается из вестибулярных ядер, проходит на той и другой стороне по бокам средней линии, непосредственно под серым веществом дна водопровода и IV желудочка, и состоит из восходящих и нисходящих волокон, идущих к ядрам III, IV, VI и XI черепных нервов.
Медиальный продольный пучок является важным ассоциативным путем, связующим различные ядра нервов глазных мышц между собой, чем обусловливаются сочетанные движения глаз при отклонении их в ту или другую сторону. Функция его связана также с движениями глаз и головы, возникающими при раздражении аппарата равновесия.
, fibrae corticonucleares. Часть пирамидного пути к двигательным ядрам черепных нервов. Рис. Б, Рис. В.
Корково-мостовые волокна
Теменно-височно-мостовые волокна
Лобно-мостовые волокна
, fibrae frontopontinae. Часть корково-мостового пути, которая начинается в лобной доле и занимает медиальную 1/6 часть ножки мозга. Рис. Б, Рис. В.
Черное вещество
, substantia nigra. Расположено в ножке мозга и определяется невооруженным глазом, т.к. состоит из пигментсодержащих нейронов. Рис. Б, Рис. В.
Компактная часть
Сетчатая часть
, pars reticularis. Вентральная часть черного вещества. Состоит из пигментсодержащих волокон и диффузно расположеных скоплений нейронов.
Покрышка среднего мозга
, tegmentum mesencephalicum. Находится между черным веществом и плоскостью, проходящей через водопровод мозга параллельно пластинке четверохолмия . Рис. Б, Рис. В.
Центральное серое вещество
Крыша среднего мозга
, tectum mesencephalicum. Расположена кзади от плоскости, проведенной через водопровод мозга. Рис. Б, Рис. В.
Пластинка крыши [[четверохолмия]]
Нижний холмик
Верхний холмик
Ручка нижнего холмика
Ручка верхнего холмика
Треугольник петли
Верхняя ножка мозжечка
, pedunculus cerebellaris superior. Содержит волокна, следущие от зубчатого ядра мозжечка к красному ядру и таламусу. Рис. А.
Водопровод среднего мозга (водопровод мозга)
, aquaeductus mesencephali (cerebri). Узкий канал внутри среднего мозга между III и IV желудочками. Рис. Б, Рис. В, Рис. Г.
Разрезы среднего мозга
Ретикулярная формация
, formatio (substantia) reticularis. Продолжение ретикулярной формации моста. Окружает водопровод мозга и состоит из диффузно расположенных нейронов, функция которых заключается в интеграции мышечной активности. Рис. Б, Рис. В, Рис. Г.
Корково-ретикулярные волокна
, fibrae corticoreticulares. Направляются от двигательных областей коры большого мозга к нейронам ретикулярной формации.
Медиальный продольный пучок
, fasciculus longitudinalis medialis. Соединяет двигательные ядра III, IV, VI, XI и вестибулярные ядра VIII черепных нервов с мотонейронами передних рогов, иннервирующих мышцы шеи. Рис. Б, Рис. В, Рис. Г.
Задний продольный [[Шютца]] пучок
, fasciculus longitudinalis posterior [[Schutz]]. Соединяет между собой гипоталамус, добавочное, верхнее и нижнее слюноотделительные, двойное ядра, заднее ядро блуждающего нерва, ядро одиночного пути, ядра лицевого и подъязычного нервов. Рис. Г.
Среднемозговой путь тройничного нерва
, tractus mesencephalicus nervi trigeminalis. Волокна V нерва, направляющиеся к одноименному ядру с латеральной стороны водопровода среднего мозга. Рис. Г.
Ядро среднемозгового пути тройничного нерва
, nucleus tractus mesen cephalicinervi trigemini (nucl. mes. trigeminalis). Расположено под пластинкой крыши. Рис. Г.
Ядро глазодвигательного нерва
, nucleus nervi oculomotorii (nucleus oculomotorius). Находится спереди от водопровода мозга. Рис. Б.
Добавочное ядро глазодвигательного нерва
, nucleus oculomotorius accessorius (autonomicus). Парасимпатическая часть глазодвигательного ядра. Большинство его клеток (96%) иннервируют ресничную мышцу, остальные - сфинктер зрачка.
Ядро блокового нерва
, nucleus nervitrochlearis (nucleus trochlearis). Находится в центральном сером веществе каудальнее ядра глазодвигательного нерва. Рис. В.
Межножковое ядро
, nucleus interpeduncularis. Расположено в области дна межножковой ямки и соединяется с обонятельным трактом. Рис. В.

Обоснование. До настоящего времени нет общепринятой схемы пространственной организации групп нейронов компактной части черного вещества (ЧВ, substantia nigra) среднего мозга человека. Детальное исследование цитоархитектоники этого образования необходимо для патоморфологического анализа изменений, происходящих в нервной ткани с возрастом, и развивающихся при этом нейродегенеративных заболеваний, сопровождающихся избирательной гибелью дофаминовых нейронов.
Цель. Уточнить особенности морфохимической организации ЧВ мозга человека и провести пространственную реконструкцию структур его компактной части.
Материалы и методы. На аутопсийном материале мозга людей без неврологической патологии (n=10, возраст от 52 до 84 лет) методом компьютерной морфометрии произвели пространственную реконструкцию компактной части ЧВ, используя срезы среднего мозга, окрашенные по Нисслю и иммуногистохимически – для локализации тирозингидроксилазы – маркера дофамина.
Результаты. Выявили в компактной части ЧВ скопления нейронов в форме 9 тяжей, ориентированных в рострокаудальном направлении, которые представили в виде 4 областей: медиальной, латеральной, дорсальной и вентральной. Морфометрический анализ обнаружил значимые различия в плотности расположения нейронов и показателях экспрессии тирозингидроксилазы между областями ЧВ.
Заключение. Схема клеточной организации компактной части ЧВ, предложенная нами на основании трехмерной реконструкции, отличается высокой детализацией по сравнению с подобными работами и демонстрирует выраженную пространственную дифференцировку групп нейронов ЧВ, что необходимо учитывать при патоморфологических исследованиях.
Ключевые слова
Полный текст
Общеизвестно, что физиологическое старение и болезнь Паркинсона (БП) проявляются морфологическими изменениями черного вещества (ЧВ) головного мозга. Вместе с тем, возрастную инволюцию ЧВ характеризует естественная убыль нейронов [1], а болезнь Паркинсона – избирательная гибель дофаминовых нейронов в его компактной части [2]. Интенсивность и однородность количественных изменений нейронов в структурах ЧВ как в условиях физиологического старения, так и в условиях патологии – БП, возможно оценить с помощью морфометрического исследования. Вместе с тем, углубленная оценка количественных изменений клеточных структур ЧВ основана на исследовании соответствующих параметров отдельных скоплений нейронов, составляющих это образование мозга. В связи с этим, выдвигались различные варианты разделения ЧВ на отдельные клеточные структуры. В одном из них ЧВ делилось на две зоны – черную (компактную часть), представленную 21 группой пигментированных нейронов, и красную (ретикулярную часть), состоящую из слабопигментированных клеток [3]. Другие авторы объединяли в 6 обособленных групп дофаминовые нейроны, расположенные в области среднего мозга [4], а дофаминовые нейроны ЧВ группировали по нигросомам – зонам иммунореактивности кальбиндина – кальцийсвязывающего белка, содержащегося в нигростриатных афферентных волокнах и клеточном нейропиле [5]. Кроме того, были предложены схемы разделения компактной части ЧВ мозга человека на области, доступные для изучения морфометрическими методами [6].
На основе подобных схем и классификаций у лабораторных животных был проведен анализ распределения холинергических [7] и дофаминовых нейронов [8] в ЧВ головного мозга, и описаны модели пространственной структурной организации ЧВ. В то же время, сведений, характеризующих подобные трехмерные модели организации нейронных структур компактной части ЧВ мозга человека, в доступной нам литературе мы не нашли.
Цель работы – уточнить особенности морфохимической организации ЧВ мозга человека и провести пространственную реконструкцию структур его компактной части.
Материалы и методы
Результаты и их обсуждение
На неокрашенных макропрепаратах – фронтальных срезах среднего мозга структуры ЧВ выявлялись в виде отдельного тяжа темнокоричневого цвета (рис. 1,а). При окрашивании по Нисслю в ЧВ различали две зоны скоплений нейронов (рис. 1,б): одна из них прилежала к ножке мозга и была более густоклеточной, другая располагалась ниже и латеральнее красного ядра. Границы ЧВ определяли по локализации дофаминовых нейронов и их отростков (рис. 1,в), а местоположение этого образования мозга по отношению к другим структурам ствола – при помощи пространственной реконструкции, проведенной по срезам среднего мозга, окрашенным на ТГ (рис. 2,а).
На полученной трехмерной модели пространственной организации ЧВ, построенной на основании срезов среднего мозга, окрашенных по Нисслю, компактная часть ЧВ (рис. 2,б) была представлена 9ю тяжами, состоящими из скоплений групп нейронов, и эти тяжи были ориентированы в рострокаудальном направлении. При проецировании построенной модели на плоскость в компактной части ЧВ выделили 4 области: медиальную, латеральную, дорсальную и вентральную (рис. 2,б).
Морфометрическое исследование плотности расположения нейронов в выделенных нами областях компактной части ЧВ показало, что плотность их расположения была выше в медиальной и вентральной области, чем в латеральной и дорсальной областях (рис. 3,а).
Оценка интенсивности иммуногистохимического окрашивания на ТГ в этих областях (за исключением латеральной области) обнаружила наибольшую экспрессию ТГ в медиальной области ЧВ (рис. 3,б).
Проведенное исследование позволило уточнить пространственную локализацию скоплений групп нейронов в ЧВ мозга людей старших возрастных групп и разграничить компактную часть ЧВ на отдельные области, доступные для изучения методами математической морфологии. Работы по трехмерной реконструкции черной субстанции мозга крыс и мышей демонстрируют сходное цитоархитектоническое деление компактной части ЧВ грызунов на три или четыре области, в зависимости от того, выделяют ли авторы медиальную область [8, 11]. Вместе с тем, повидимому, группы дофаминовых нейронов ЧВ грызунов являются менее дифференцированными, по сравнению с человеком, что выражается в отсутствии четко различимого сегментарного разделения отделов ЧВ и подтверждается как настоящей работой, так и другими авторами [12].

Рис. 1. Локализация черного вещества на поперечном срезе среднего мозга человека. а – макропрепарат, фиксация формалином; б – окраска методом Ниссля; в – иммуногистохимическое выявление тирозингидроксилазы. ЧВ – черное вещество; 1,2 – вентральная, редкоклеточная, и дорсальная, густоклеточная, области, соответственно; пунктир с точкой – границы компактной части ЧВ

Рис. 2. Объемная реконструкция черного вещества среднего мозга человека. а – границы и относительное расположение в среднем мозге дофаминергических структур черной субстанции по локализации тирозингидроксилазы (синим); б – объемная организация нейрональных скоплений в каудальных 2/3 черного вещества, на основе окрашивания крезиловым фиолетовым. ЧВ – черное вещество; НМ – ножка мозга; КЯ – красное ядро. Группы нейронов черной субстанции (1-9): медиальная область – 1,2; дорсальная область – 6-8; вентральная область – 3-5; латеральная область – 9

Рис. 3. Количественные показатели областей черной субстанции. а – плотность распределения нейронов на 0,1 мм3; б – распределение интенсивности окрашивания (в условных единицах яркости) на тирозингидроксилазу в каудо-ростральном направлении (по оси абсцисс – номера срезов через 0,2 мм). Д – дорсальная область; В – вентральная область; М – медиальная область; Л — латеральная область
Согласно результатам нашего исследования, компактная часть ЧВ головного мозга человека состоит из 9 тяжей (представляющих собой скопления групп нейронов), которые при сопоставлении с данными литературы [13] соответствуют следующим сегментам: вентромедиальному – тяжи 1 и 2, промежуточному – тяж 3, вентролатеральному – тяжи 4 и 5, дорсомедиальному – тяж 6, дорсолатеральному – тяжи 7 и 8. Латеральной части дорсальной области соответствует тяж 9. Предложенная нами пространственная модель ЧВ головного мозга показывает неполное соответствие при сравнении с другими вариантами организации ЧВ [14, 15], что выражается выявленной нами большей детализацией структур этого образования мозга. Такое несоответствие, возможно, обусловлено тем, что один из упомянутых выше вариантов разделения ЧВ предполагал изучение зон иммунореактивности кальбиндина, и оба варианта – не предполагали применение методов трехмерной реконструкции.
Морфометрическое исследование компактной части ЧВ показало, что плотность расположения нейронов в медиальной области была выше, чем в других областях компактной части ЧВ (за исключением вентральной), и интенсивность окрашивания на ТГ в медиальной области была наибольшей, что соответствует результатам других исследователей [13]. В связи с этим, можно предположить, что у лиц старших возрастных групп инволютивные изменения в меньшей степени затрагивают медиальную область этого образования головного мозга. По данным литературы, разные группы дофаминовых нейронов среднего мозга отличаются не только локализацией и своими связями, но и различной уязвимостью к действию повреждающих факторов в эксперименте, при нейродегенеративной патологии и при физиологическом старении [16]. Причины этого до конца не выяснены, но предполагают, что в основе избирательной устойчивости отдельных групп дофаминовых нейронов лежат различия их нейрохимического профиля, обусловливающие их подверженность окислительному стрессу [17].
Заключение
Таким образом, пространственная структурная морфохимическая организация черного вещества головного мозга человека отличается тем, что на всем его протяжении выявляли скопления нейронов, входящих в состав 9 тяжей, ориентированных в рострокаудальном направлении, которые при проецировании на горизонтальную плоскость формировали 4 области: медиальную, латеральную, дорсальную и вентральную. Морфометрическое исследование этих областей позволило обнаружить, что медиальная область черного вещества оказалась менее подверженной возрастной инволюции.
Читайте также: