
Двойное оплодотворение характерно для простейших покрытосеменных грибов
Обновлено: 07.07.2024
У покрытосеменных, цветковых растений уникальный процесс оплодотворения. Именно уникальный. Ни один другой живой организм не проходит через процесс, который биологи называют «Двойное Оплодотворение«.
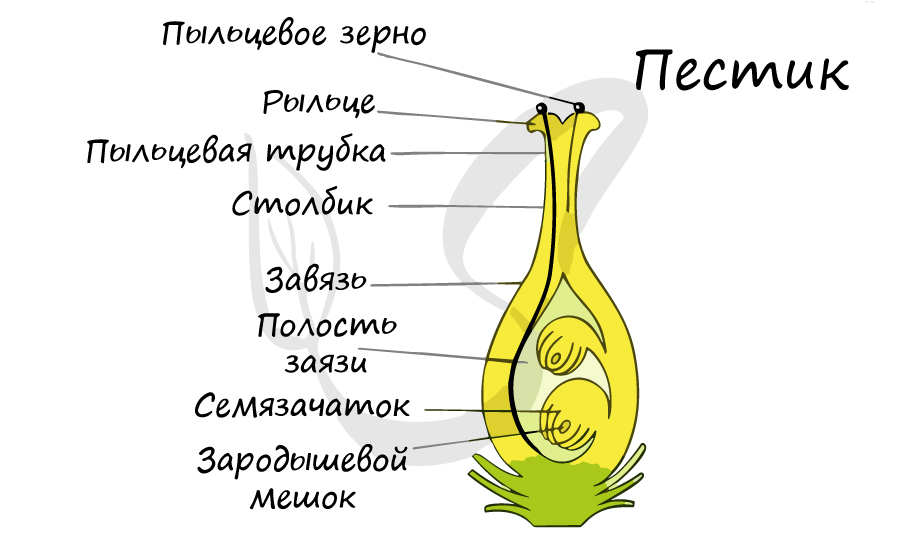
На женском гаметофите в семязачатке развивается мегаспора — женская половая клетка, которая формируется в завязи пестика.
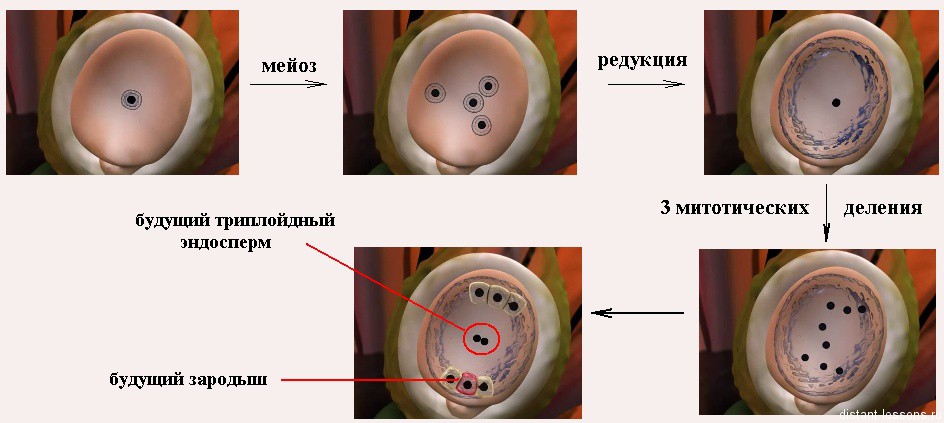
Спорогенная клетка (2n) сначала делится митозом. Образуются мегаспороциты (2n). Затем они делятся мейозом, образуются гаплоидные (1n) клетки — мегаспоры. У большинства видов 3 из этих клеток редуцируются и остается одна мегаспора.
Она развивается и ее ядро 3 раза делится митозом. Образуются 8 гаплоидных ядра, которые находятся в цитоплазме одной клетки. 6 ядер смещаются к краям семязачатка, 2 ядра остаются в центре клетки.
- Два слившихся ядра образуют центральное ядро,
- остающиеся на полюсах ядра превращаются в клетки:
— антиподы (1n),
— яйцеклетку (1n),
— синергиды (1n) - формируется женский гаметофит, или зародышевый мешок

Мужские половые клетки — микроспоры.
В состав пыльцевого зерна входят 2 клетки:
- вегетативная клетка, которая в будущем формирует пыльцевую трубку и
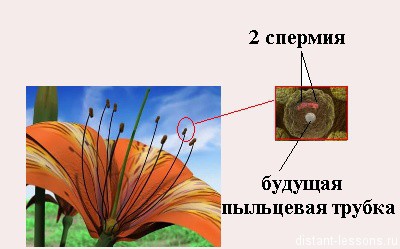
- генеративная клетка, которая в будущем митозом образует 2 спермия.
Опыление — процесс переноса пыльцы с пыльников тычинок цветка на рыльце пестика
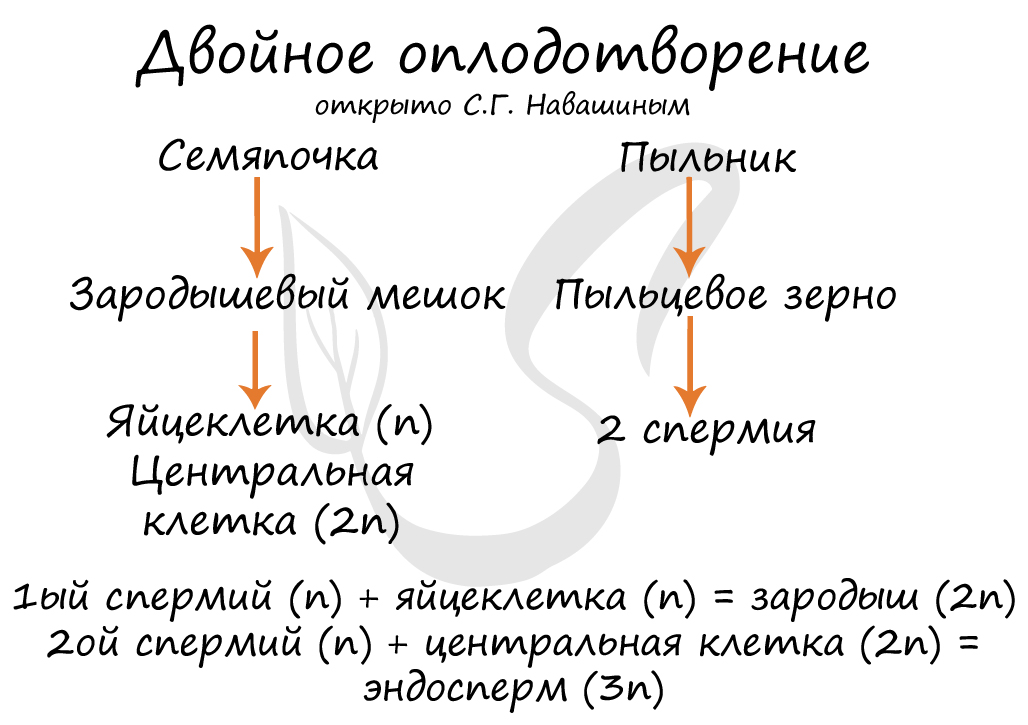
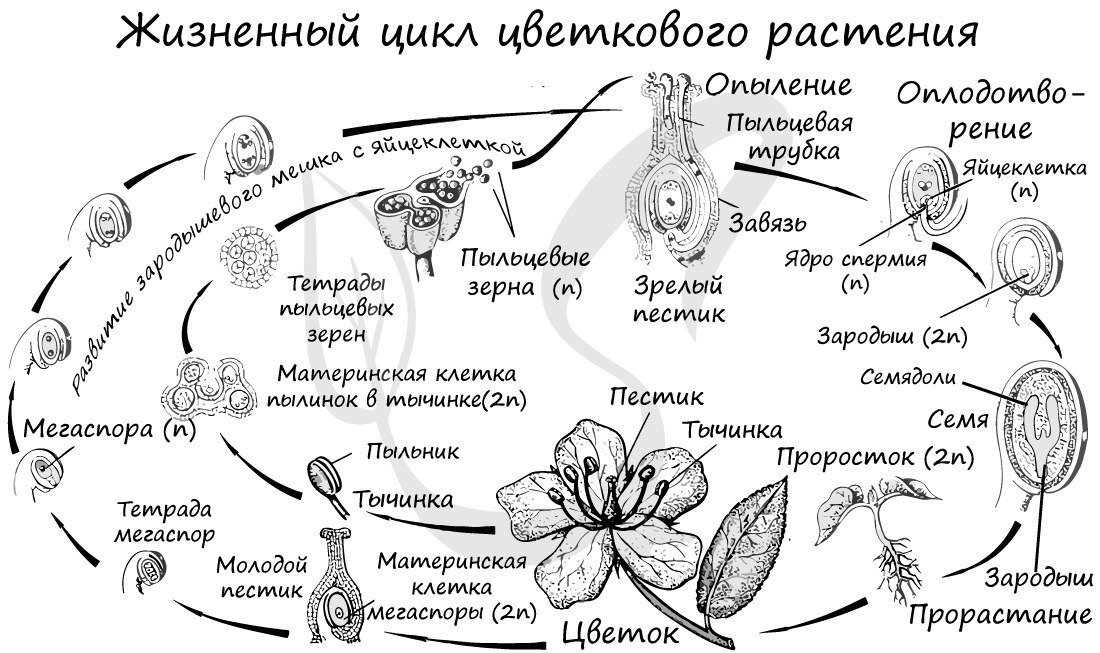
Пыльцевой зерно попадает на рыльце пестика, образующаяся пыльцевая трубка начинает прорастать внутрь столбика, стремясь к завязи. Когда пыльцевая трубка достигает зародышевого мешка, генеративная клетка образует 2 спермия. Один из спермиев оплодотворяет яйцеклетку, образуется зародыш. Второй спермий сливается с двумя ядрами в середине, образуя триплойдную клетку (3n) — эндосперм. Это и есть двойное оплодотворение.
Процесс двойного оплодотворения был открыт С.Г. Навашиным в 1898 г
Последовательность этапов этого процесса можно свести к схеме:
(такой вопрос часто встречается в части B ЕГЭ)

- после оплодотворения семязачаток превращается в семя
- завязь превращается в плод,
- из зиготы — зародыш семени
- из покровов семязачатка формируется семенная кожура.
Значение двойного оплодотворения
В семенах очень быстро (опережая развитие зародыша) образуются питательные ткани с запасом высокоэнергетических питательных веществ только после оплодотворения. У цветковых растений при развитии семязачатков не тратится время на создание питательных веществ, как у голосеменных растений, поэтому развиваются они гораздо быстрее. Таким образом, благодаря двойному оплодотворению ускоряется процесс формирования и семяпочки, и яйцеклетки, и семени
У голосеменных в оплодотворении участвует один спермий и эндосперм возникает до оплодотворения в результате деления мегаспоры, т.е. он гаплоидный и первичный.
Среди цветковых растений широко распространен апомиксис — образование зародыша без оплодотворения ( без участия мужской гаметы). У животных есть аналогичный процесс — партеногенез. Зародыш может развиваться из любых клеток зародышевого мешка: яйцеклетки, центрального ядра, синергид или антипод. Из остальных частей семязачатка образуются другие части семени.
Двудомные растения опыляются извне. Соответственно, это либо опыление животными (насекомыми, в основном), либо ветром.
Для привлечения насекомых растения используют яркую окраску цветка и аромат, пыльца крупная, липкая; ветроопыляемые растения обычно имеют невзрачные цветки, которые высоко поднимаются над землей и мелкую сухую пыльцу.
половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка (См. Зародышевый мешок). Д. о. открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений — лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия (См. Спермии) сливается с ядром яйцеклетки, ядро второго — с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается Зародыш, из центральной клетки — Эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид (См. Синергиды), которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой — проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При Д. о. ядра зародышевого мешка находятся в интерфазе (См. Интерфаза) и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа Д. о.: премитотическое — ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое — мужское и женское ядра, сохраняя свои оболочки, вступают в профазу (См. Профаза), в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности (См. Плоидность); у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие Д. о. — Ксении — проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в Д. о., спермий остальных — дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Лит.: Навашин С. Г., Избр. труды, т. 1, М.— Л., 1951; Магешвар и П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Поддубная Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Steffen К., Fertilisation, в кн.: Maheshwari P. (ed.). Recent advances in the embryology of angiosperms, Delhi, 1963.
Двойное оплодотворение; 1 — у рябчика: один из спермиев (а) в контакте с ядром яйцеклетки, второй (б) — с одним из полярных ядер (второе полярное ядро не изображено); 2 — у подсолнечника: а — пыльцевая трубка; б — синергиды (одна из них повреждена пыльцевой трубкой); в — яйцеклетка; г — спермий в контакте с ядром яйцеклетки; д — центральная клетка; е — второй спермий в контакте со вторичным ядром зародышевого мешка.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру: от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.

Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) - важнейшими производителями органических веществ на суше, как водоросли - в морях и океанах.
Цветок - генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения. Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище - завязи, сформированной из плодолистика (-ов).
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества - эндосперма (3n). Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод - генеративный орган, служащий для защиты и распространения семян.
Ксилема - проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами, а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

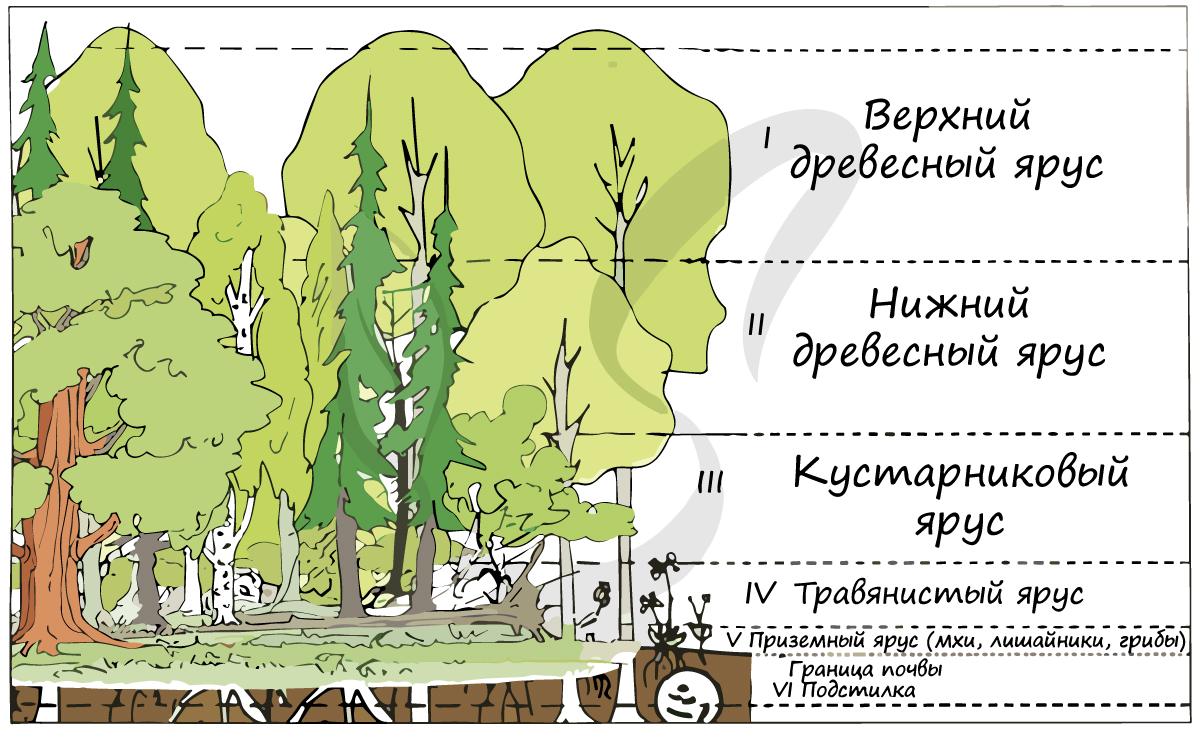
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых :)
Классы покрытосеменных
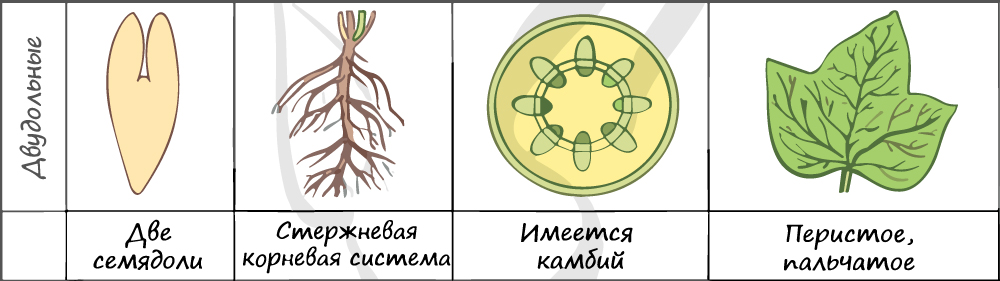
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные, бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
-
Двудольные - семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
-
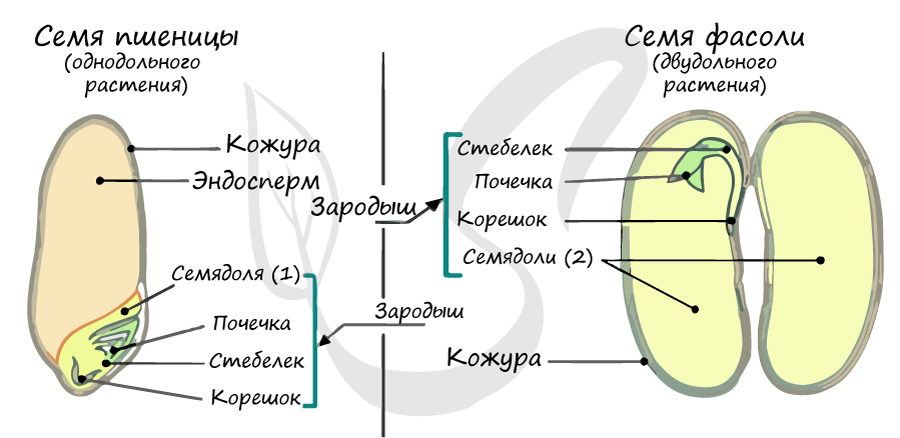
В составе зародыша обычно имеется две семядоли
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни. Главный корень развивается из зародышевого корешка.
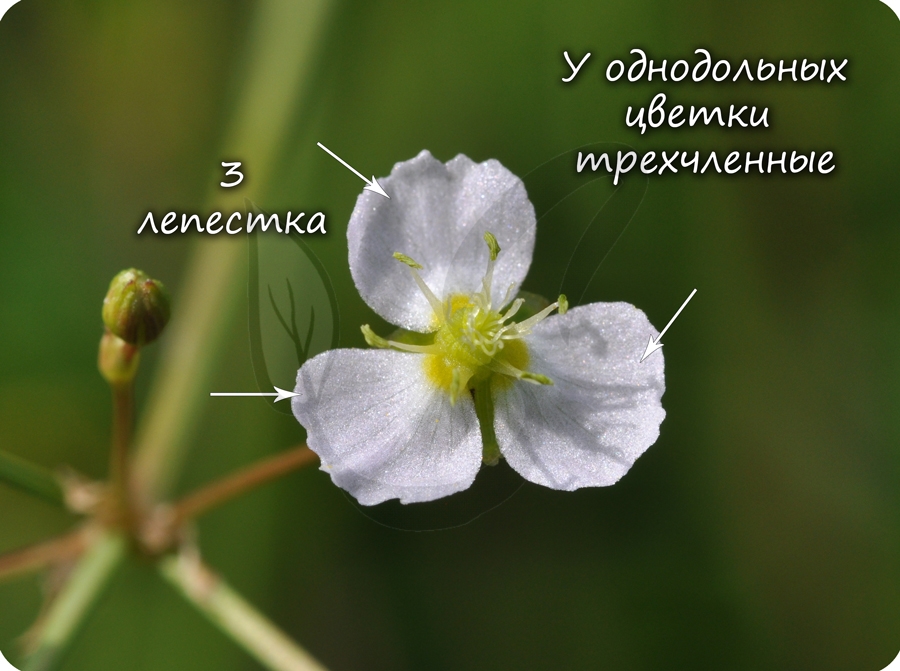
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

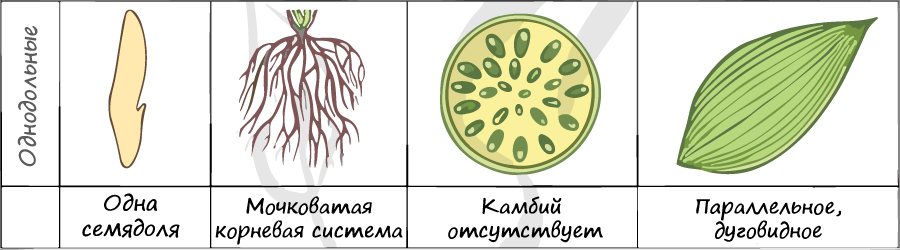
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
Эндосперм семени
Эндосперм (от греч. endon - внутри + греч. sperma - семя) - запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у - березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
Жизненный цикл
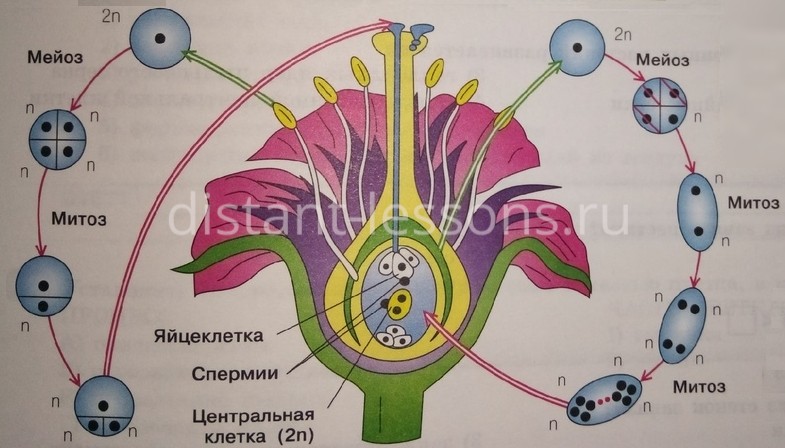
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n) в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) - мужской гаметофит. В завязи пестика в семязачатке формируется женский гаметофит - зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток. Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) - запасного питательного вещества. Другой спермий (n) сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник - образуется плод. Своим внешним видом плоды привлекают животных, и те их охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия, они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте - почти все культурные растения принадлежат к этому отделу! Цветковые имеют медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.

Видеоурок способствует формированию представлений о способах опыления цветковых растений, выявляет приспособления к различным способам опыления. Основная цель урока – рассмотреть особенности двойного оплодотворения и его преимущества. В ходе видеоурока подробно рассказывается о процессах микроспорогенеза и макроспорогенеза, их сущности.

В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Двойное оплодотворение покрытосеменных растений"
Размножение – это одно из обязательных свойств любого живого организма. Оно заключается в увеличении числа особей.
Различают бесполое и половое размножение растений.
Бесполое размножение подразделяют на спорообразование и вегетативное.
Половое размножение происходит при помощи особых половых клеток –гамет.
При бесполом размножении быстро увеличивается численность вида, все потомки имеют абсолютно такой же генотип, что и родительская особь. А также не происходит увеличения генетического разнообразия, которое может оказаться очень полезным при изменении условий существования вида.
По этой причине большинство живых организмов на Земле размножаются половым путём.
Сущность полового размножения заключается в слиянии генетической информации родителей, благодаря чему генетическое разнообразие в потомстве увеличивается.
У покрытосеменных растений половое размножение связано с цветком.
Важные части цветка – это пестик и тычинка. С их участием происходят сложные процессы полового размножения –опыление и оплодотворение.
Но сперва в будущем цветке начинают образовываться половые клетки.
В жизненном цикле цветковых растений наблюдается смена поколений.
У растений выделяют диплоидное поколение – бесполое, или спорофит, и гаплоидное поколение – половое, или гаметофит.
Гаметы образуются в результате митоза, а споры – в результате мейоза. И гаметы, и споры образуются в цветке, поэтому цветок является органом и бесполого, и полового размножения.
Пыльцевое зерно является спорой (микроспорой), а не мужской гаметой, так как в нем самом развиваются мужские гаметы.
У цветковых растений мужские гаметофиты столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего лишь из нескольких клеток.
Женский гаметофит цветковых (зародышевый мешок) помещается внутри семяпочки и состоит в наиболее распространённом случае из 7 клеток (содержит 8 – либо 7 после слияния двух ядер в центральной клетке и образования вторичного ядра).
Посмотрим, как образуется мужской гаметофит.
Итак, Микроспорогенез
В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делений становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
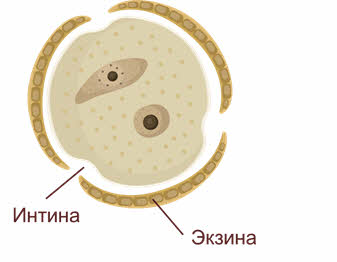
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, поверхность её либо гладкая, либо шероховатая, приспособленная для переноса пыльцы и прилипания её к рыльцу пестика.
Этим заканчивается микроспорогенез, вслед за образованием одноядерной микроспоры начинается микрогаметогенез.
Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и её ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться ещё в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, называются спермиоклетками, или спермиями.
Таким образом, из одной споры (микроспоры) с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: два из них – спермии и одно – вегетативное. При образовании пыльцевой трубки это вегетативное ядро переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены российским и советским цитологом и эмбриологом растений Сергеем Гавриловичем Навашиным в 1910 г. на лилейных растениях.
После образования гамет пыльник созревает, и пыльца высыпается. Она несёт только генетическую информацию.
Посмотрим, как происходит мегаспорогенез и мегагаметогенез цветковых.
У покрытосеменных растений женский гаметофит – это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез.
В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растёт, превращаясь в материнскую клетку мегаспоры.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют, судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем её ядро претерпевает ряд делений. При этом сама клетка не делится, а делится только ядро.
У разных систематических групп растений число делений ядра мегаспоры может варьировать от одного до трёх. У большинства растений (70 % видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других – в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
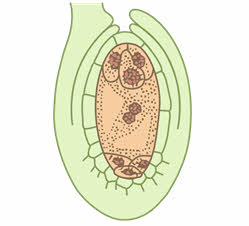
В дальнейшем от каждой из двух полярных четвёрки ядер к центру отходит по одному ядру, которые сливаются, образуя вторичное (центральное) ядро зародышевого мешка. Затем цитоплазма обособляется вокруг ядер гаметофита, который из ядерной стадии развития переходит в клеточную. Три ядра, оставшиеся вблизи халазального полюса, преобразуются в три клетки (антиподы), которые питают гаметофит. Три ядра вблизи микропиле отделяются клеточными перегородками, образуя отдельные клетки: крупную центральную яйцеклетку и две боковые клетки синергиды. Вся цитоплазма, расположенная между антиподами с одной стороны и клетками яйцевого комплекса с другой (яйцеклетка и две синергиды), называется центральной клеткой. В ней находится диплоидное вторичное (центральное) ядро.
На этом этапе женский гаметофит уже полностью сформирован и состоит из шести гаплоидных клеток (одной яйцеклетки, двух синергид и трёх антипод) и одной диплоидной (центральной). Его строение внешне напоминает мешочек, поэтому женский гаметофит покрытосеменных называется зародышевым мешком.
Пылинка попадает на рыльце пестика, и происходит опыление.
Опыление – это перенос пыльцевых зёрен на рыльце пестика, у голосеменных пыльцевые зерна при опылении попадают непосредственно на семязачаток.
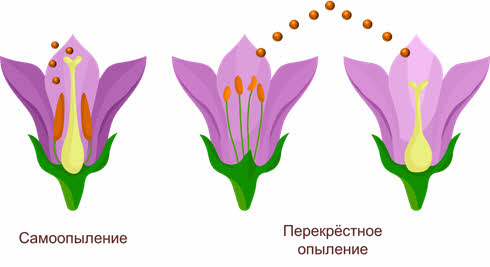
Имеется два основных типа опыления: самоопыление (автогамия) (когда растение опыляется собственной пыльцой) и перекрёстное опыление (аллогамия).
При самоопылении исключён обмен генетической информацией, поскольку пыльцевые зерна попадают на рыльце пестика либо с одной из тычинок этого же цветка, либо с другого цветка, расположенного на том же растении. Это приводит к появлению чистых линий гомозиготных популяций в пределах одного вида, неспособных обмениваться мутировавшими генами, поэтому процессы видообразования в этих популяциях идут самостоятельно.
Перекрёстное опыление – это перенос пыльцы одного растения на рыльце другого. Этот тип опыления встречается более часто, чем самоопыление, между разными особями одного вида происходит обмен аллелями, что приводит к увеличению доли гетерозиготных организмов.
Безусловно, перекрёстное самоопыление имеет большие преимущества по сравнению с самоопылением, поскольку возникшие мутации свободно распространяются в пределах популяции.
Способы опыления у перекрёстноопыляемых цветковых растений весьма разнообразны. Их можно разделить на две группы. Первая: перенос пыльцы осуществляется главным образом насекомыми, а также некоторыми позвоночными (птицами и летучими мышами). Соответственно, различают энтомофилию, орнитофилию и зоофилию. Растения, опыляемые животными, обычно имеют яркоокрашенные крупные цветки. Мелкие цветки, как правило, собраны в соцветия, что зрительно их увеличивает. Для привлечения опылителей служит нектар или большое количество пыльцы, которую опылители охотно поедают.
Вторая группа: пыльца переносится абиотическими факторами – ветром и реже водой, в связи с чем различают анемофилию и гидрофилию.
После попадания пыльцы на рыльце начинается прорастание пыльцевого зерна.
Экзина мужского гаметофита прорывается в области борозды или поры прорастания, и начинает формироваться пыльцевая трубка, которая растёт, проникает в рыльце, через столбик движется по направлению к завязи, где находится семязачаток.
Трубка растёт на кончике. В растущую пыльцевую трубку из пыльцевого зерна перемещается ядро клетки-трубки, а также спермин.
Обычно развитие пыльцевой трубки происходит при уже сформированных семязачатках в завязи. Достигнув завязи, пыльцевая трубка через микропиле проникает внутрь одного из находящихся там семязачатков.
Там пыльцевая трубка направляется к яйцевому аппарату, проникает в одну из синергид и освобождает спермин. Один из спермиев сливается с яйцеклеткой, в результате чего образуется зигота.
Другой спермий сливается с полярными ядрами центральной клетки, образуя триплоидное ядро (с тройным набором хромосом). Зигота даёт начало зародышу, а из триплоидной центральной клетки образуется ткань эндосперма.
Таким образом оплодотворение, при котором одна мужская гамета сливается с яйцеклеткой, а вторая ― с вторичным ядром, называется двойным оплодотворением.
Механизм оплодотворения цветковых –двойное оплодотворение, открыто отечественным учёным Сергеем Гавриловичем Навашиным в 1898 году.
Таким образом, оплодотворённый семязачаток развивается в семя, из зиготы возникает зародыш, из триплоидной центральной клетки – эндосперм, а из внешней части семязачатка (интегументов) образуется семенная кожура, из стенок завязи цветка – стенки плода.
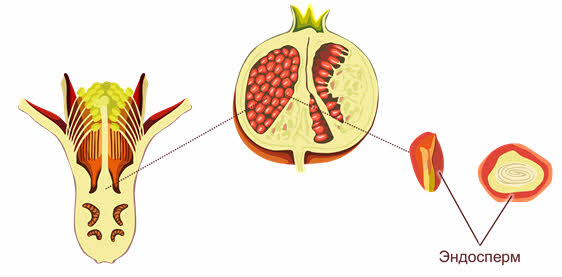
Эндосперм цветковых полностью отличается от первичного эндосперма голосеменных. У них первичный эндосперм представляет собой гаплоидную вегетативную ткань женского гаметофита, где накапливаются питательные вещества семени.
Эндосперм у покрытосеменных происходит из триплоидной центральной клетки, в образовании которой участвуют вторичное ядро женского гаметофита и ядро спермия.
У одних цветковых (например, злаков) эндосперм сильно разрастается и занимает большую часть семени, оттесняя на периферию маленький зародыш.
У других (к пример у бобовых) весь эндосперм поглощается зародышем и используется его семядолями, которые становятся самыми большими структурами зрелого семени.
Суть двойного оплодотворения у цветковых растений заключается в том, что в нём участвуют два спермия. Один из них оплодотворяет яйцеклетку, и образуется зигота. Второй спермий сливается с центральной клеткой, из которой развивается запасающая ткань (эндосперм).

Мужской гаметофит (пыльцевое зерно) образуется в пыльцевых камерах пыльников тычинки из микроспоры. Пыльцевое зерно состоит из двух гаплоидных клеток: вегетативной и генеративной, покрытых оболочкой.
Образование женского гаметофита (зародышевого мешка) происходит в завязи пестика в семязачатке из мегаспоры. В состав зародышевого мешка входит семь клеток: гаплоидная яйцеклетка, центральная диплоидная клетка и пять вспомогательных гаплоидных клеток.
При попадании пыльцевого зерна на рыльце пестика начинается деление вегетативной клетки и образуется пыльцевая трубка. Пыльцевая трубка прорастает через столбик пестика и проникает в семязачаток через пыльцевход.
Генеративная клетка пыльцевого зерна делится и образует два спермия. По пыльцевой трубке спермии проникают в семязачаток. Один спермий сливается с яйцеклеткой и образует диплоидную зиготу. Второй спермий сливается с центральной клеткой и образует триплоидную клетку.
Зигота делится, и развивается в зародыш нового растения. Из триплоидной клетки формируется эндосперм. Стенки семязачатка становятся семенной кожурой. Таким образом, семязачаток становится семенем.
Читайте также: