
Физиология цветения опыления и оплодотворения
Обновлено: 07.07.2024
Цветение растений — одно из самых привлекательных явлений природы, приводящее к образованию плодов и семян.
Цветок — это видоизмененный побег с укороченным стеблем, листья которого превратились и в цельные части. Цветок обычно сидит на цветоножке, Он состоит из зеленых чашелистиков, которые пли срастаются между собой (шалфей, будра), или свободны (лютик, пастушья сумка). Чашечка защищает венчик, иногда видоизменяется в летучку и служит для разноса плодов (одуванчик, осот, бодяк и др.).
Венчик состоит из лепестков, окрашенных в разные цвета. Форма венчика чрезвычайно разнообразна, главное его назначение — привлекать споим цветом, запахом и нектаром опылителей.
Органами размножения являются тычинки и пестик. Каждая тычинка имеет тычиночную нить и пыльник. Пыльник состоит из двух половинок, соединенных связником. К связнику прикрепляется тычиночная нить. Пыльца у вётроопыляемых растений мелкая и сухая, а у опыляемых насекомыми крупная и липкая. Если тычинок не более 12, то, говоря о растении, указывают их число, если больше 12, то отмечают, что растения многотычинковые.
Число пестиков также различно. Пестик состоит из нижней части (завязи), средней (столбика) и верхней (рыльца).
Растение, у которого пестичные и тычиночные цветки находятся на одном и том же экземпляре, называют однодомным (огурец, тыква, дуб, береза и др.). Если мужские и женские цветки находятся на разных растениях, то их называют двудомными (конопля, ива, осина и др.). У бесполых часть цветков утрачивает тычинки и пестики (василек, подсолнечник).
Мелкие цветки часто бывают собраны в соцветия разной формы: кисть (черемуха), щиток (боярышник), метелка (овес), сережка (береза), колос (пшеница), початок (кукуруза), зонтик (морковь, петрушка), головка (клевер), корзинка (подсолнечник).
Опыление — это перенос пыльцы с тычинок на пестик. Если пыльца с тычинок переносится на свой же пестик, то происходит самоопыление (фиалка, яснотка, звездчатка, горох, соя, пшеница, ячмень, овес, просо и др.). Если пыльца с тычинок одного растения переносится на пестик другого, то происходит перекрестное опыление.
Большая часть растений опыляется перекрестно, причем и основном за счет насекомых и только десятая часть растений — ветром. Цветки имеют ряд приспособлений для опыления насекомыми: яркую окраску и сильный запах, которые должны привлекать опылителей, а в награду опылители получают пыльцу и сладкий нектар. Насекомые прихватывают пыльцу с собой на лапках; брюшке и переносят па другое растение, производя тем самым процесс опыления. Если нектар спрятан, неглубоко, то растения опыляют мухи, жуки. Если нектар спрятан глубоко, то его могут достать насекомые с длинным язычком (пчелы, шмели). Иногда у растений (табак) цветки имеют длинные трубки, в которые проникают только длинные хоботки бабочек.
Если стоит дождливая погода, то насекомые мало летают и не переносят пыльцу, образуется много пустоцветов — неопыленных растений.
У ветроопыляемых растений (рожь, кукуруза) цветки невзрачны, в них не образуется нектара, зато тычинки имеют длинные нити, которые, свешиваясь, позволяют ветру беспрепятственно переносить, пыльцу. Пыльцы производится много, и при таком малоэффективном способе опыления ее очень много пропадает.
У растений имеются различные приспособления, которые обеспечивают перекрестное опыление, позволяющее избежать самоопыления. У ветроопыляемых растений цветки часто раздельнополые; у растений, опыляемых насекомыми, в большинстве случаев тычинки созревают быстрее, чем формируется пестик. У медуницы, незабудки, дербенника цветки имеют неодинаковые по длине тычинки и пестики.
Оплодотворение. Пыльца, попавшая на рыльце пестика, начинает прорастать. Образовавшаяся при этом пыльцевая трубка растет по направлению к семяпочке (достигая иногда в длину 2 см, например у кукурузы). Конец пыльцевой трубки разрывается, и оттуда выходят две мужские клетки. Одна из них сливается с женской яйцеклеткой, а другая - со вторичным ядром зародышевого мешка. Происходит двойное оплодотворение. Оплодотворенная клетка несет в себе наследственные признаки мужского и женского растения. Семяпочка превращается в семя, а завязь - в плод. Оплодотворенное вторичное ядро путем деления образует запасную вторичную ткань, необходимую для развития зародыша.
Плоды и семена. В образовавшемся зародыше возникают семядоли: у однодольных — одна, у двудольных — две. Образуется также корешок, подсемядольное колено и почечка. Питательные вещества находятся в семядолях.
Процесс созревания плодов и семян у разных растений протекает за разное время. У раннецветущих плоды созревают быстро (за 2—3 недели), у многих растений — за 2—3 месяца.
Плоды подразделяют на сухие и сочные. Сухие бывают односемянные (нераскрывающиеся), например семянка, орех, зерновка, и многосемянные (раскрывающиеся), например боб, стручок, коробочка. У сочных плодов выделяют костянку (вишни, слива, персик), ягоду (помидор, виноград, крыжовник), сложные плоды (малина, земляника), ягодообразные (арбуз, огурец, яблоко, груша).
Созревшие плоды и семена распространяются различными способами, широко расселяясь на поверхности земли.
Водой переносятся плоды многих водных растений (осока, белая кувшинка). Особенно интересны плоды кокосовой пальмы, которые океан выбрасывает на сушу, нередко очень далеко от места их созревания.
Растения-катапульты выработали ряд приспособлений, позволяющих им выбрасывать семена далеко от материнского растения. К ним относятся сухие плоды бобовых (боб), кислица, недотрога, бешеный огурец. Клетки таких плодов находятся под большим давлением, стоит к ним прикоснуться, как давление изменяется, створки сильно закручиваются, далеко выбрасывая семена.
Ряд растений имеет особые приспособления: самозарываться за счет штопорообразных остей (ковыль, овсюг), зацепляться за животных при помощи крючков (лопух, репейник, череда).
Распространение животными и человеком. Некоторые животные заготавливают себе па зиму съестные припасы, но часть их не съедают, давая растениям возможность распространяться (желуди, орехи, шишки). Птицы, поедая сочные плоды, способствуют распространению многих растений (вишня, боярышник, рябина, малина и др.). Человек распространяет семена диких растений при перевозке и при плохой обработке посевного материала.
Процессы, связанные с оплодотворением, разделяют на четыре фазы: опыление, прорастание пыльцы на рыльце, рост пыльцевых трубок в тканях столбика и собственно оплодотворение.
Опыление. В зрелом состоянии мужской и женский гаметофиты имеют минимальную физиологическую активность. При попадании пыльцы на рыльце пестика в зоне соприкосновения происходят активные метаболические процессы. Пыльца начинает поглощать воду из рыльца и набухать. При этом она выделяет на поверхность рыльца белки, АК, НК, углеводы, липиды, пигменты, гидролитические ферменты и др. ФАВ.
Начальная фаза адгезии (слипания) происходит вследствие поступления подвижных веществ на поверхность рыльца из внешней оболочки пыльцы — экзины, затем начинают высвобождаться вещества из интины.
С. И. Лебедевым (1982) установлена особая роль в оплодотворении каротиноидов, которые обусловливают желтый цвет андроцея и гинецея, как правило, энтомофильных растений. Под влиянием рассеянного света такая пыльца приобретает запах фиалки или земляники вследствие освобождения из каротина летучего вещества (3-ионона), чем и привлекаются насекомые.
Проникновение (инвазия) желтых пигментов пыльцы в ткани рыльца активизирует прорастание пыльцевых зерен. При прорастании пыльцы ксантофиллы играют роль аккумуляторов кислорода для ОВР. Наряду с влиянием каротиноидов на прорастание пыльцы их недостаток приводит к нарушению микроспорогенеза и образованию абортивных гамет. Каротиноиды также принимают активное участие в двойном оплодотворении (В. А. Поддубная-Арнольди, 1976).
Пестик и его рыльце обладают менее высоким содержанием и активностью ферментов и других физиологических веществ, чем пыльца, тем не менее гинецей играет важную роль в процессе прорастания пыльцы.
Выделения рылец влажного типа представлены липофильными экссудатами и гидрофильными слизистыми секретами, насыщенными белками и углеводами. Эти выделения содержат также гидролитические ферменты (пероксидазы, эстеразы, амилазы). Выделяемые рыльцем секреторные вещества обеспечивают адгезию пыльцы, поступление в нее воды при прорастании, защиту от инфекции, поддерживают рост пыльцевой трубки в тканях рыльца.
Рост пыльцевой трубки в столбике пестика. В зависимости от анатомической структуры столбики разделяют на два основных типа- открытый и закрытый. Открытый тип (характерен для однодольных растений) имеет специальный канал, выполненный простой эпидермой и физиологически активной слизью. Через этот канал происходит рост пыльцевой трубки. У закрытого типа столбика (характерен для двудольных растений) канал заполнен проводящей тканью, через которую осуществляется врастание трубки в завязь.
Пыльцевая трубка изначально образуется из протопласта пыльцевого зерна, вовлекая в себя вегетативное ядро и генеративную клетку, а в дальнейшем используются питательные вещества, содержащиеся в проводниковой ткани столбика пестика. Рост пыльцевой трубки сопровождается повышенной активностью метаболических процессов: увеличивается интенсивность дыхания, изменяется водный режим, наблюдается усиленное образование ауксина и триптофана.
За счет активного действия гидролитических ферментов растворяются пектиновые вещества, связывающие клетки проводящих тканей, что облегчает проникновение трубки через столбик. В физиологической среде в области роста пыльцевых трубок увеличивается содержание углеводов, белков, фосфорных соединений, аскорбиновой кислоты, ауксинов, гиббереллинов и других веществ.
Процесс роста пыльцевых трубок предопределен наличием в пыльце рибосомальной и информационной РНК. Во время этого процесса новый синтез РНК не наблюдается. Из минеральных веществ особо активное участие принимают ионы кальция, калия, бора, магния и др.
У большинства покрытосеменных растений при прорастании пыльцевой трубки происходит образование спермиев путем митотического деления генеративной клетки. Однако у многих покрытосеменных растений сперматогенез происходит в пыльниках до наступления цветения.
Оплодотворение. Пыльцевая трубка проникает через микропиле в зародышевый мешок, где она входит в контакт с одной из синергид. Содержимое трубки, ее цитоплазма, вегетативное ядро и спермии сливаются с цитоплазмой синергиды. Один из спермиев оплодотворяет яйцеклетку, в результате чего образуется диплоидная зигота, дающая начало зародышу. Другой спермии сливается с диплоидным вторичным ядром зародышевого мешка, что приводит к возникновению триплоидной первичной клетки эндосперма. В этом заключается свойственное покрытосеменным растениям двойное оплодотворение, открытое С. Г. Навашиным в 1898 г.
При излиянии содержимого пыльцевой трубки в зародышевый мешок в нем создается определенная физиологическая среда, которая в случае совместимости женских и мужских половых элементов будет способствовать процессу оплодотворения, а при несовместимости — препятствовать ему.
Ч. Дарвин впервые описал самонесовместимость при самоопылении перекрестно-опыляемых растений. Она заключается в способности отторгать собственную пыльцу из-за разницы в тургорном давлении в пыльце и рыльце либо вследствие ингибирования прорастания образованием полисахарида каллозы.
Известно, что на начальных этапах прорастания пыльцевого зерна из его оболочек (экзины и интины) на поверхность рыльца пестика начинают выделяться специфические белки, которые у совместимого рыльца вызывают активацию физиолого-биохимических процессов, способствующих образованию и росту пыльцевой трубки, а у несовместимого — образование каллозы, изолирующей пыльцевое зерно и таким образом вызывающей прекращение его прорастания. У некоторых растений в клетках несовместимого пестика синтезируются белки, являющиеся ингибиторами роста пыльцевых трубок.
Кроме физиологических и биохимических причин самонесовместимости она может обусловливаться морфологическими особенностями цветка. Примером может служить гетеростилия — наличие цветков с различной длиной столбиков пестиков и нитей тычинок у разных растений одного и того же вида, что препятствует самоопылению.
В физиологическом смысле цветение – это комплекс процессов, протекающих в период от заложения цветка до оплодотворения.
Процесс цветения делят на две фазы: I) инициацию заложения цветочных зачатков; 2) развитие из зачатков цветков вплоть до их раскрывания.
Цветок может возникнуть из верхушечной меристемы главного или боковых побегов или из тех и других. Когда растение достигает этапа половой зрелости, некоторые или все апикальные меристемы на его побегах перестают закладывать листья и начинают формировать части цветка.
В клетках меристемы, готовой к заложению цветка, усиливается синтез нуклеиновых кислот, увеличивается количество рибосом и белков. Кроме того, изменяется состав синтезируемых белков, повышается активность некоторых ферментов. Одновременно увеличивается количество митохондрий, дыхательных субстратов и усиливается дыхание меристематических клеток. В меристеме увеличивается количество митозов и образуются новые клетки, дающие начало цветку.
Сначала увеличивается конус нарастания, затем он превращается в цветковый (флоральный) бугорок. В его меристеме первоначально выделяются три зоны: периферическая, средняя и центральная. Из периферической зоны формируются элементы околоцветника, из средней – андроцей, а из центральной – гинецей. У большинства видов двудольных растений первыми закладываются чашелистики или листочки околоцветника, затем пыльники. В результате выделяется центральная часть, превращающаяся в пестик, последними закладываются лепестки. Апикальная меристема после образования всех частей цветка теряет свою активность. Зачатки органов одного круга цветка всегда возникают одновременно.
Затем заложенные органы начинают расти. Первыми растут чашелистики, защищающие органы цветка, потом тычиночные нити, пестик и лепестки. Постепенно лепестки приобретают окраску, соответствующую виду. Окраска лепестков обусловлена каротиноидами (пигментами хромопластов), антоцианами (пигментами клеточного сока) и зависит от кислотности клеточного сока.
В ранний период своего формирования цветок является актиноморфным. У видов, имеющих зигоморфные или асимметричные цветки, они возникают из актиноморфных в результате последующего неравномерного роста элементов околоцветника.
В случае образования соцветия клетки меристемы побега, быстро делясь и растягиваясь, образуют сначала ось соцветия, затем начинается заложение цветков.
В пыльниках происходит микроспорогенез, образуются микроспоры. Из каждой микроспоры формируется мужской гаметофит – пыльца, состоящая из двух клеток – генеративной и клетки трубки – и покрытая двумя оболочками – экзиной и интиной.
В завязи образуется семязачаток, состоящий из нуцеллуса, двух интегументов и семяножки (фуникулуса). В семязачатке происходит мегаспорогенез и образуются четыре мегаспоры. Одна из них делится и из нее развивается женский гаметофит, называемый зародышевым мешком и содержащий гаплоидную яйцеклетку, синергиды, антиподы и диплоидное вторичное (центральное) ядро. У большинства растений к моменту опыления все элементы женского гаметофита сформированы.
Пыльца и ткани пестика имеют разные свойства, влияющие на активность ферментов, химические реакции и физиологическое состояние. Особенно большие различия в значениях pH: у пыльцы рН>7, у рыльца рН
Перед опылением зрелая пыльца и зрелый пестик имеют минимальную физиологическую активность. При попадании пыльцы на рыльце пестика происходит их слипание (адгезия) и начинаются очень активные метаболические процессы, активное взаимодействие пыльцы и рыльца. Такое рыльце называют воспринимающим. Пыльца выделяет на поверхность рыльца белки, аминокислоты, нуклеиновые кислоты, углеводы, липиды, пигменты, гидролитические ферменты и другие физиологически активные вещества. Каротиноиды, находящиеся в пыльце, проникают в ткани рыльца и активизируют прорастание пыльцы. Из рыльца в пыльцу поступает вода, и она набухает. Перед прорастанием пыльцы метаболизм в тканях рыльца и столбика менее активен, чем у пылинки.
Пыльца, попав на рыльце, прорастает. Инициируют прорастание пыльцы поступление воды и активизация синтеза ферментов. При набухании пылинки экзина лопается или вскрывается в определенных местах, а интина вытягивается в пыльцевую трубку, в которую переходит генеративная клетка. Вскоре генеративная клетка делится, и из нее образуются два спермия. У некоторых растений спермии возникают, когда пыльца находится в пыльнике. В конце пыльцевой трубки располагаются вегетативное ядро, аппарат Гольджи и эндоплазматический ретикулум, участвующие в синтезе веществ для растущей клеточной стенки.
Пыльца хорошо прорастает только на рыльцах растений своего вида; на рыльцах растений других видов пыльца или не прорастает совсем, или пыльцевая трубка растет плохо и не может достичь завязи. У многих растений, требующих обязательно перекрестного опыления, например у плодовых деревьев, пыльца плохо прорастает на рыльце своего цветка. Предполагают, что в цветке находятся вещества, тормозящие прорастание своей пыльцы. Это явление получило название физиологической самостерильности растений.
Пыльца может сохранять способность к прорастанию в течение некоторого времени, продолжительность которого зависит от вида растения и условий ее хранения. У большинства злаков пыльца сохраняет способность к прорастанию лишь в течение 1-3 дней, у некоторых орхидных – 178 дней, а у финиковой пальмы – целый год.
Прорастание пыльцы зависит от температуры, поэтому холода во время цветения часто неблагоприятно влияют на урожайность. Вредное действие оказывают дожди и туманы, так как не только погибает пыльца, но с рыльца смываются сахара и другие необходимые для прорастания вещества. Поэтому цветки перед дождем закрываются.
Пыльцевая трубка сначала внедряется в ткань рыльца, а затем проникает в столбик и растет в нем все глубже и глубже, направляясь к завязи. Столбики делят на два типа: сплошные или полые с каналом в центре. Эти каналы выстланы проводниковой тканью, выделяющей слизь и служащей источником питательных веществ для растущей пыльцевой трубки. По этому каналу и растет пыльцевая трубка. Такие столбики характерны для однодольных растений. Сплошной столбик характерен для двудольных. Он не имеет канала, внутри заполнен проводниковой тканью, клетки которой расположены вертикальными рядами и погружены в основную ткань. Пыльцевая трубка достигает проводниковой ткани и растет вдоль нее. Растущий конец пыльцевой трубки выделяет ферменты, способные растворять пектиновые вещества, связывающие клетки проводниковой ткани, что облегчает рост пыльцевой трубки в столбике.
Пыльцевая трубка обладает сильно выраженным хемотропизмом, что помогает ей выбирать правильное направление для роста. Направленный рост пыльцевой трубки определяется возрастанием концентрации ионов кальция. Концентрация кальция увеличивается от рыльца по длине столбика и далее к завязи; еще большая концентрация в семязачатке, а наивысшая – в зародышевом мешке. Необходимые для роста пыльцевой трубки вещества уже заранее отложены в пыльце. Пыльца богата питательными веществами.
Рост пыльцевой трубки сопровождается увеличением интенсивности дыхания, изменением водного обмена, усиленным образованием ауксина и триптофана. Около растущей пыльцевой трубки увеличивается содержание сахаров, белков, фосфорных соединений, аскорбиновой кислоты, ауксинов, гиббереллинов и других веществ, а также ионов кальция, калия, бора, магния.
Минимальное время, необходимое для достижения пыльцевой трубкой семяпочки, равно 15 мин. Чаще всего этот срок составляет несколько часов, а иногда еще больше. Например, у березы он длится целый месяц, у ольхи и лещины – от месяца до двух, а у некоторых орхидных – 6-7 мес. Достигнув завязи, пыльцевая трубка растет вдоль ее внутренней стенки и попадает в семязачаток, как правило, через микропиле.
Оплодотворение.Пыльцевая трубка подходит к зародышевому мешку. Здесь она проникает в одну из синергид и разрывается, вероятно, под действием ферментов, выделяемых зародышевым мешком. Попав в зародышевый мешок, один спермий соединяется с яйцеклеткой, образуя зиготу, а второй спермий – со вторичным ядром, образуя триплоидное ядро. Происходит двойное оплодотворение.
Зигота обычно некоторое время находится в покое. После двойного оплодотворения начинается развитие зародыша и формирование семени. Одновременно завязь увеличивается в размерах и из нее развивается плод. Околоцветник и тычинки обычно засыхают и опадают, столбик после опыления у большинства видов тоже засыхает.

Опыление и оплодотворение растений. Половое размножение
Половое размножение характерно для большинства растений, за исключением некоторых водорослей.
Опыление — это перенос пыльцы с тычинок на рыльце пестика. Различают перекрестное опыление и самоопыление (рис.1).

Рис.1 Виды опыления цветковых растений
При самоопылении происходит перенос пыльцы с тычинок на рыльце пестика в пределах одного и того же цветка.
В природе самоопыление происходит редко, зачастую еще в бутонах, до раскрытия цветка (пшеница, горох, фасоль, фиалка, томат). Главное преимущество самоопыления — оно не зависит от погодных условий и насекомых, поэтому осуществляется при любых условиях. Не все обоеполые цветки являются самоопыляющимися. Большинство растений дают полноценные семена только при перекрестном опылении.
Опыление, при котором пыльца тычинок одного цветка попадает на рыльце пестика другого, называют перекрестным. Перекрестное опыление осуществляется насекомыми и ветром. Реже — птицами, летучими мышами и водой.
Строение цветков насекомоопыляемых растений разнообразно (вишня, слива, яблоня, сирень, роза и многие другие). Они имеют ярко окрашенный или белый венчик и сильный запах. Цветки крупные или собраны в соцветия.
Запах цветков и их яркая окраска привлекают насекомых. Пчелы, шмели, мухи, бабочки, жуки и муравьи питаются пыльцой и нектаром цветка. Нектарники, расположенные в глубине цветка, выделяют нектар до тех пор, пока цветок не завянет. Тело насекомого, пытающегося добраться до нектарников, обильно покрывается пыльцой. Перелетая с одного цветка на другой в поисках пищи, они переносят прилипшую к их телу пыльцу с тычинок одних цветков на рыльца пестиков других.
Опыление ветром возникло в процессе эволюции как приспособление к неблагоприятным условиям. Надежда на немногочисленных тогда мух, бабочек, пчел и других насекомых была слабой. Позже насекомых стало больше. Но наряду с насекомоопыляемыми растениями, существуют опыляемые ветром. Это многие злаковые травы лугов, степей и саванн, обитатели леса (береза, ольха, осина, дуб, орешник), пустынь и полупустынь (полыни, солянки) (рис.2).
У ветроопыляемых растений бывает очень много пыльцы. Она легкая, сухая и мелкая. Околоцветник отсутствует или плохо развит и не препятствует движению ветра. Перистые рыльца пестиков приспособлены к улавливанию пыльцы. Тычиночные нити длинные и свисающие. Раскачиваясь на ветру, они распыляют зрелую пыльцу.
Большинство ветроопыляемых растений цветет до распускания листьев, что облегчает опыление. Но оно зачастую зависит от погоды. В облачные, дождливые дни осадки смывают пыльцу и тем самым снижают урожай.
Искусственное опыление осуществляет человек, перенося пыльцу с тычинок на рыльца пестиков. Такое опыление требует больших затрат времени и трудно осуществимо на больших площадях. Чаще всего к нему прибегают при выведении новых сортов.
Оплодотворение (рис.2) происходит после опыления.Пыльца, или пыльцевое зерно, попав на рыльце пестика, выделяющего липкую жидкость, прорастает, образуя длинную, тонкую пыльцевую трубку. Пыльцевая трубка, постепенно удлиняясь, продвигается по столбику рыльца по направлению к завязи (нижняя, важнейшая часть пестика). В завязи образуются семязачатки (семяпочки). Снаружи они защищены покровами, а внутри находится зародышевый мешок, состоящий из нескольких клеток.
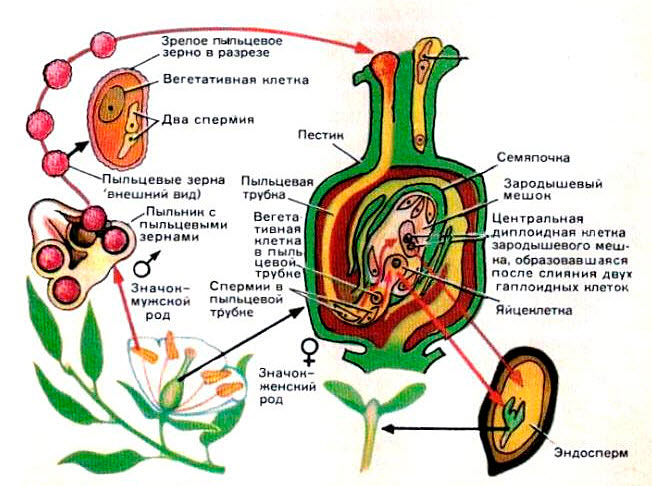
Рис.2 Процесс двойного оплодотворения
Одна из клеток в зародышевом мешке — яйцеклетка, это женская половая клетка (женская гамета). Другая — крупная центральная клетка.
В пыльцевой трубке находятся две маленькие мужские половые клетки (мужские гаметы) — спермии. Когда пыльцевая трубка достигает семязачатка и врастает в него, один спермий сливается с яйцеклеткой. Слияние двух половых клеток (гамет) называется оплодотворением. Из оплодотворенной яйцеклетки впоследствии разовьется зародыш растения. Второй спермий сливается с крупной центральной клеткой. Из нее образуется ткань эндосперм. В клетках этой ткани накапливаются питательные вещества для развития зародыша. Покровы семязачатка превращаются в семенную кожуру. Таким образом два одинаковых спермия сливаются с двумя разными женскими гаметами. Происходит двойное оплодотворение (открыл у лилейных русский ученый в 1898 г. Г.Навашин). После оплодотворения образуется семя, которое состоит из зародыша, запасающей ткани (эндосперма) и семенной кожуры. Из стенки завязи образуется плод.
Опыление — перенос пыльцы на рыльце пестика. Бывает самоопыление — внутри закрытого цветка (горох, пшеница) и перекрестное: насекомыми — цветки яркие, с запахом (яблоня, сирень, роза); ветром — цветки без околоцветника, распускаются до появления листьев, много мелкой легкой пыльцы (береза, дуб, полынь). Искусственное опыление проводит человек. После опыления происходит оплодотворение — процесс слияние мужских и женских половых клеток — гамет. Двойное оплодотворение — два спермия из пыльцы, достигнув завязи пестика по пыльцевой трубке, оплодотворяют две женские гаметы. При слиянии яйцеклетки и спермия образуется зародыш. При слиянии второго спермия и центральной клетки образуется эндосперм (запас питательных веществ). Из покровов семязачатка образуется кожура семени, а из стенки завязи пестика — плод.
Читайте также: