
Какими веществами прокариотическая водоросль может снабжать микобионт в лишайнике
Обновлено: 05.10.2024
Лишайники — своеобразная группа живых организмов, произрастающих на всех континентах, в том числе и в Антарктиде. В природе их насчитывают более 26 000 видов.
Долгое время лишайники были загадкой для исследователей. Однако до сих пор не пришли к единому мнению относительно их положению в систематике живой природы: одни относят их к царству растений, другие — к царству грибов.
Тело лишайника представлено слоевищем. Оно очень разнообразно по окраске, размерам, форме и строению. Слоевище может иметь форму тела в виде корочки, листовидной пластинки, трубочек, кустика и небольшого округлого комочка. Некоторые лишайники достигают в длину более метра, но большинство имеют слоевище размером 3-7 см. Они медленно растут — за год увеличиваются на считанные миллиметры, а некоторые — на доли миллиметра. Возраст их слоевища нередко насчитывает несколько сотен и тысяч лет.
Лишайники не имеют типичной зелёной окраски. Окраска лишайников сероватая, зеленовато-серая, светло- или тёмно-бурая, реже жёлтая, оранжевая, белая, чёрная. Окраска обусловлена пигментами, которые находятся в оболочках гиф гриба. Различают пять групп пигментов: зелёные, синие, фиолетовые, красные, коричневые. Цвет лишайников может зависеть также от окраски лишайниковых кислот, которые откладываются в виде кристаллов или зёрен на поверхности гиф.
Живые и отмершие лишайники, скопившаяся на них пыль и песчинки создают не обнажённом грунте тонкий слой почвы, в котором могут закрепиться мхи и другие наземные растения. Разрастаясь, мхи и травы затеняют наземные лишайники, засыпают их отмершими частями своих тел, и лишайники со временем исчезают с этого места. Лишайникам вертикальных поверхностей засыпание не грозит — они разрастаются и разрастаются, впитывая влагу дождей, рос и туманов.
В зависимости от внешнего облика слоевища лишайники делят на три типа: накипные, листоватые и кустистые.
Типы лишайников. Морфологические особенности
Лишайники — первые поселенцы на обнажённом грунте. На голых камнях, палимых солнцем, на песке, на брёвнах и стволах деревьев.
(около 80% всех лишайников)
Вид корочки, тонкой плёнки, разных цветов тесно сросшихся с субстратом
В зависимости от субстрата, на котором произрастают накипные лишайники, различают:
- эпилитные
- эпифлеоидные
- эпигейные
- эпиксильные
на поверхности горных пород;
на коре деревьев и кустарников;
на поверхности почвы;
на гниющей древесине
Слоевище лишайника может развиваться внутри субстрата (камня, коры, дерева). Есть накипные лишайники с шаровидной формой слоевища (кочующие лишайники)
Таллом имеет вид чешуек или достаточно больших пластинок.
Монофильное — вид одной крупной округлой листовидной пластинки (в диаметре 10—20 см).
Полифильное — слоевище из нескольких листовидных пластинок
Прикрепляются к субстрату в нескольких местах с помощью пучков грибных гиф
На камнях, почве, песке, коре деревьев. К субстрату прочно прикрепляются толстой короткой ножкой.
Встречаются неприкреплённые, кочующие формы
Характерной особенностью листовидных лишайников является то, что его верхняя поверхность отличается по строению и окраске от нижней
Кустистые.
Высота маленьких — несколько миллиметров, крупных — 30—50 см
Слоевища бывают с плоскими и округлыми лопастями. Иногда у крупных кустистых лишайников в условиях тундр и высокогорий развиваются добавочные прикрепительные органы (гаптеры), с помощью которых они прирастают к листьям осок, злаков, кустарников. Таким образом, лишайники предохраняют себя от отрыва сильными ветрами и бурями
Эпифиты — на ветвях деревьев или скалах. К субстрату прикрепляются небольшими участками слоевища.
Напочвенные — нитевидными ризоидами
Уснея длинная — 7—8 метров, свисающая в виде бороды с ветвей лиственниц и кедров в таёжных лесах
Это высший этап развития слоевища
В чрезвычайно суровых условиях произрастают лишайники на камнях и скалах в Антарктиде. Живым организмам приходится жить здесь при очень низких температурах, особенно зимой, и практически без воды. Из-за низкой температуры осадки там выпадают всегда в виде снега. Лишайник не может поглощать воду в такой форме. Но его выручает чёрная окраска слоевище. Благодаря высокой солнечной радиации тёмная поверхность тела лишайника быстро нагревается даже при низких температурах. Снег, попавший на нагретое слоевище, тает. Появившуюся влагу лишайник сразу впитывает, обеспечивая себя водой, необходимой ему для дыхания и фотосинтеза.
Строение
Слоевище состоит из двух разных организмов — гриба и водоросли. Они так тесно взаимодействуют между собой, что их симбиоз представляется единым организмом.
Слоевище представляет собой множество переплетённых грибных нитей (гиф).

Между ними группами или одиночно расположены клетки зелёных водорослей, а у некоторых — цианобактерий. Интересно, что виды грибов, составляющих лишайник, в природе вообще не существуют без водорослей, тогда как большинство водорослей, входящих в слоевище лишайника, встречается в свободноживущем состоянии, отдельно от гриба.

Питание
Питание лишайника осуществляется обоими симбионтами. Гифы гриба поглощают воду и растворённые в ней минеральные вещества, а водоросль (или цианобактерия), в которой имеется хлорофилл, образует органические вещества (благодаря фотосинтезу).
Гифы играют роль корней: они впитывают воду и растворённые в ней минеральные соли. Клетки водорослей образуют органические вещества, выполняют функцию листьев. Воду лишайники впитывают всей поверхностью тела (используют дождевую воду, влагу туманов). Важным компонентом в питании лишайников является азот. Те лишайники, которые в качестве фикобионта имеют зелёные водоросли, получают соединения азота из водных растворов, когда их слоевище пропитывается водой, частично прямо из субстрата. Лишайники, имеющие в качестве фикобионта сине-зелёные водоросли (особенно ностоки), способны фиксировать атмосферный азот.
Внутреннее строение
Это своеобразная группа низших растений, которые состоят из двух разных организмов — гриба (представители аскомицетов, базидиомицетов, фикомицетов) и водоросли (зелёные — цистококк, хлорококк, хлорелла, встречается кладофора, пальмелла; сине-зелёные — носток, глеокапса, хроококк), образующих симбиотическое сожительство, отличающееся особыми морфологическими типами и особыми физиолого-биохимическими процессами.
По анатомическому строению различают лишайники двух типов. У одного из них водоросли разбросаны по всей толще слоевища и погружены в слизь, которую выделяет водоросль (гомеомерный тип). Это наиболее примитивный тип. Такое строение характерно для тех лишайников, фикобионтом которых являются сине-зелёные водоросли. Они образуют группу слизистых лишайников. У других (гетеромерный тип) на поперечном срезе можно под микроскопом различать несколько слоёв.

Сверху находится верхняя кора, имеющая вид переплетённых, тесно сомкнутых грибных гиф. Под ней гифы лежат более рыхло, между ними расположены водоросли — это гонидиальный слой. Ниже грибные гифы расположены ещё более рыхло, большие промежутки между ними заполнены воздухом — это сердцевина. За сердцевиной следует нижняя кора, которая по строению подобна верхней. Через нижнюю кору из сердцевина проходят пучки гиф, которые прикрепляют лишайник к субстрату. У корковых лишайников нижней коры нет и грибные гифы сердцевины срастаются непосредственно с субстратом.
У кустистых радиально построенных лишайников на периферии поперечного разреза находится кора, под ней гонидиальный слой, а внутри — сердцевина. Кора выполняет защитную и укрепляющую функции. На нижнем коровом слое лишайников обычно образуются органы прикрепления. Иногда они имеют вид тонких нитей, состоящих из одного ряда клеток. Их называют ризоидами. Ризоиды могут соединяться, образуя ризоидальные тяжи.
У некоторых листовых лишайников слоевище прикрепляется с помощью короткой ножки (гомфа) расположенной в центральной части слоевища.
Зона водорослей выполняет функцию фотосинтеза и накопления органических веществ. Основная функция сердцевина — проведение воздуха к клеткам водорослей, содержащим хлорофилл. У некоторых кустистых лишайников сердцевина выполняет и укрепляющую функцию.
Органами газообмена служат псевдоцифеллы (разрывы коры, заметные невооружённым глазом как белые пятнышки неправильной формы). На нижней поверхности листовых лишайников есть круглые правильной формы белые углубления — это цифеллы, также органы газообмена. Газообмен осуществляется и через перфорации (отмершие участки корового слоя), трещины и разрывы в коровом слое.
Размножение
Размножаются лишайники главным образом кусочками слоевища, а также особыми группами клеток гриба и водоросли, во множестве образующимися внутри его тела. Под давлением их разросшейся массы тело лишайника разрывается, группы клеток разносятся ветром и дождевыми потоками. Кроме того, грибы и водоросли сохранили и свои собственные способы размножения. Грибы образуют споры, водоросли размножаются вегетативным путём.
Лишайники размножаются либо спорами, которые образуют микобионт половым или бесполым путём, либо вегетативно — фрагментами слоевища, соредиями и изидиями.
При половом размножении на слоевищах лишайников формируются половые спороношения в виде плодовых тел. Среди плодовых тел у лишайников различают апотеции (открытые плодовые тела в виде дисковидных образований); перитеции (закрытые плодовые тела, имеющие вид маленького кувшина с отверстием наверху); гастеротеции (узкие плодовые тела удлинённой формы). Большинство лишайников (свыше 250 родов) формируют апотеции. В этих плодовых телах споры развиваются внутри сумок (мешковидных образований) или экзогенно, на вершине удлинённо-булавовидных гиф — базидий. Развитие и созревание плодового тела длится 4-10 лет, а затем в течение ряда лет плодовое тело способно продуцировать споры. Спор образуется очень много: так, один апотеций может продуцировать 124 000 спор. Прорастают они не все. Для прорастания нужны условия, прежде всего определённые температура и влажность.
Бесполое спороношение лишайников — конидии, пикноконидии и стилоспоры, возникающие экзогенно на поверхности конидиеносцев. Конидии образуются на конидиеносцах, развивающихся непосредственно на поверхности слоевища, а пикноконидии и стилоспоры — в особых вместилищах пикнидиях.
Вегетативное размножение осуществляется кустиками слоевища, а также особыми вегетативными образованиями — соредиями (пылинки — микроскопические клубочки, состоящие из одной или нескольких клеток водорослей, окружённых гифами гриба, образуют мелкозернистую или порошкообразную беловатую, желтоватую массу) и изидиями (маленькие разнообразной формы выросты верхней поверхности слоевища, одного с ней цвета, имеют вид бородавочек, зёрнышек, булавовидных выростов, иногда маленьких листочков).
Лишайники — пионеры растительности. Поселяясь на местах, где другие растения произрастать не могут (например, на скалах), они через некоторое время, частично отмирая, образуют небольшое количество гумуса, на котором могут поселиться другие растения. Лишайники разрушают горные породы, выделяя лишайниковую кислоту. Это разрушительное действие заканчивают вода и ветер. Лишайники способны накапливать радиоактивные вещества.
Водоросли, встречающиеся в слоевище лишайников, называют фикобионтом лишайников. По своему систематическому положению они относятся к различным отделам: к сине-зеленым (Cyanophyta), зеленым (Chlorophyta), желто-зеленым (Xanthophyta) и бурым (Phaeophyta) водорослям. Долгое время считали, что каждому виду лишайника соответствует определенный вид водоросли. Однако, как показали дальнейшие исследования, сравнительно небольшое количество водорослей оказались способными существовать в симбиозе с грибом. Только самые неприхотливые из них, обладающие высокой устойчивостью по отношению к факторам внешней среды, приспособились к жизни в окружении грибных гиф. Экспериментальные исследования показали, что лишайниковый гриб при образовании слоевища, как правило, не проявляет избирательной способности по отношению к водорослям. Если взять чистую культуру гриба, выделенного из слоевища лишайника, и поместить в нее какую-либо свободноживущую водоросль (несвойственную данному лишайнику, а другого типа), то грибные гифы начнут разрастаться, давая сложные переплетения, как в начале образования слоевища. Однако слоевище в таком случае все-таки не развивается. Большинство водорослей, введенных в культуру лишайникового гриба, рано или поздно погибает: многие не могут перенести воздействия гиф гриба, причиной гибели других является недостаток света. Водоросли, окруженные со всех сторон грибной тканью, получают гораздо меньше света, чем это необходимо для их нормальной жизнедеятельности и если водоросль не может приспособиться к осуществлению процесса фотосинтеза при минимальной световой интенсивности, она неизбежно погибает. Значит, при образовании слоевища лишайников не гриб выбирает себе подходящего партнера из числа водорослей. Все зависит от того, сможет ли водоросль существовать в тесно.м контакте с грибом и при этом нормально жить и развиваться.
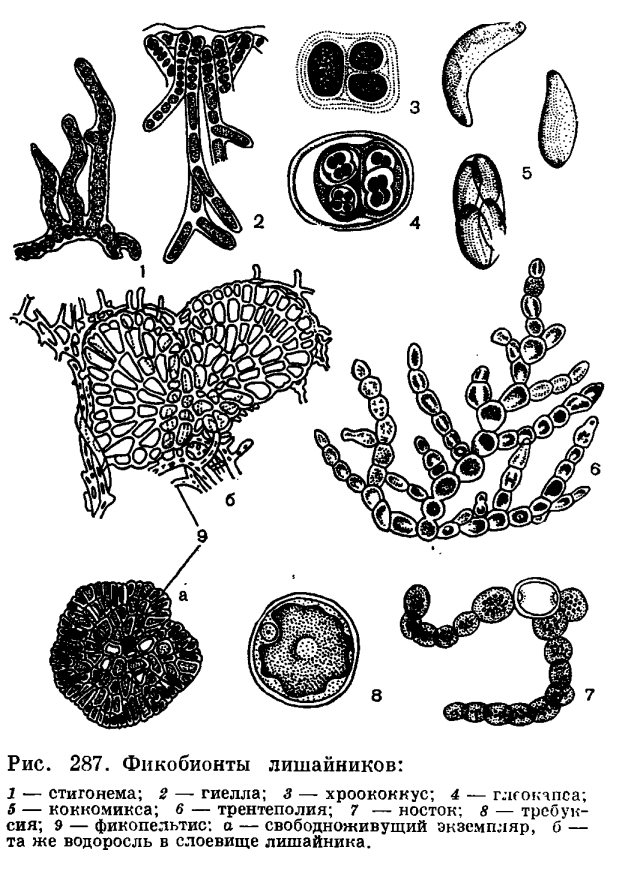
Сейчас стало известно, что наиболее широко распространенным фикобионтом лишайников является водоросль требуксия (Trebouxia, рис. 287, 8). Предполагают, что около половины всех известных видов лишайников (7 —10 тыс. видов) имеют своим фикобионтом именно эту одноклеточную зеленую водоросль. Как показали наблюдения за жизнью этой водоросли в лаборатории, требуксия очень неприхотливый организм. В условиях чистой культуры эти водоросли отличались медленным ростом и способностью развиваться при минимальном освещении, они хорошо переносили резкие перепады температуры и длительные периоды без воды. Клетки требуксии могли развиваться и в полной темноте, но в таком случае они переходили к гетеротрофному питанию. По-видимому, все эти свойства, т. е. высокая устойчивость к влиянию факторов окружающей среды, позволили требуксии так хорошо приспособиться к жизни внутри слоевища лишайников.
Как показало изучение лишайниковых фикобионтов, проведенное в последние годы, в слоевище лишайников встречаются представители 28 родов водорослей. Из сине-зеленых водорослей наиболее обычным фикобионтом в слоевище лишайников являются водоросли рода носток (Nostoc, рис. 287, 7; 288, 1) и глеокапса (Gloeocapsa, рис. 287, 4). Встречаются и другие сине-зеленые: хроококкус (Chroococcus) и гиелла (Hyella, рис. 287, 2, 3), калотрикс (Calothrix), дихотрикс (Dichothrix) и ривулярия (Rivularia, рис. 288, 2), сцитонема (Scytonema) и стигонема (Stigonema). Из зеленых водорослей в слоевище лишайников встречаются представители хлорококковых и улотриксовых. Из хлорококковых водорослей, как уже указывалось, самым распространенным фикобионтом является требуксия (Trebouxia).
Кроме того, встречается также мирмеция (Myrmecia), псевдохлорелла (Pseudochlorella), хлорококкум (Chlorococcum), хлорелла (Chlorella), коккомикса (Соссотуха, рис. 287, 5), глеоцистис (Gloeocystis) и трохисция (Trochiscia). Из улотриксовых в слоевищах лишайников были найдены представители 10 родов: цефалейрос (Cephaleuros), хлоросарцина (Chlorosarcina), коккоботрис (Coccobotrys), лептозира (Leptosira), фикопельтис (Phycopeltis, рис. 287, 9), физолинум (Physolinum), плеврококкус (Pleurococcus), псевдоплеврококкус (Pseudopleurococcus), стихококкус (Stichococcus) и трентеполия (Trentepohlia, рис. 287, 6). Желто-зеленые водоросли очень редко встречаются в лишайниковых слоевищах. Лишь у двух видов лишайников, представителей рода веррукария (Verrucaria), фикобионтом является желто-зеленая водоросль гетерококкус (Heterococcus). Так же редки в слоевищах лишайников и бурые водоросли. В слоевище одного из видов того же рода веррукария была найдена бурая водоросль петродерма (Petroderma).
Некоторые роды лишайников очень однозначны по отношению к водорослям. Так, например, все виды рода пармелия (Parmelia), широко распространенных листоватых лишайников, и кустистые лишайники рода кладония (Cladonia) имеют фикобионтом требуксию, а слизистые лишайники семейства коллемовых — носток. Но иногда у одного и того же вида лишайников бывают обнаружены разные виды водорослей. Например, у лишайника артопирения (Arthopyrenia kelpii) фикобионтом могут быть водоросли гиелла, глеокапса, псевдоплеврококкус; у листоватого лишайника солорина (Solorina saccata) в слоевище были найдены три вида рода коккомикса.
Существует взаимосвязь между географическим распространением лишайников и преобладанием в их слоевищах определенных водорослей. Так, в умеренной зоне земного шара около 8% лишайников имеют фикобионтом'сине-зеленые водоросли, 9% лишайников — нитчатые или пластинчатые зеленые водоросли типа трентеполии и 83% лишайников — хлорококковые зеленые водоросли. В тропических и субтропических странах увеличивается количество лишайников, в слоевище которых встречается водоросль трентеполия. Так, в тропиках от 5 до 10% лишайников содержат в слоевищах сине-зеленые водоросли. 45—48% лишайников — нитчатые зеленые водоросли типа трентеполии и такое же количество лишайников имеют фикобионтом остальные зеленые водоросли.
Большинство фикобионтов лишайника встречается в свободноживущем состоянии. Но некоторые известны лишь в слоевищах лишайников и до сих пор не были обнаружены в природе. К их числу, например, относятся требуксия, коккомикса, лобококкус и некоторые другие.
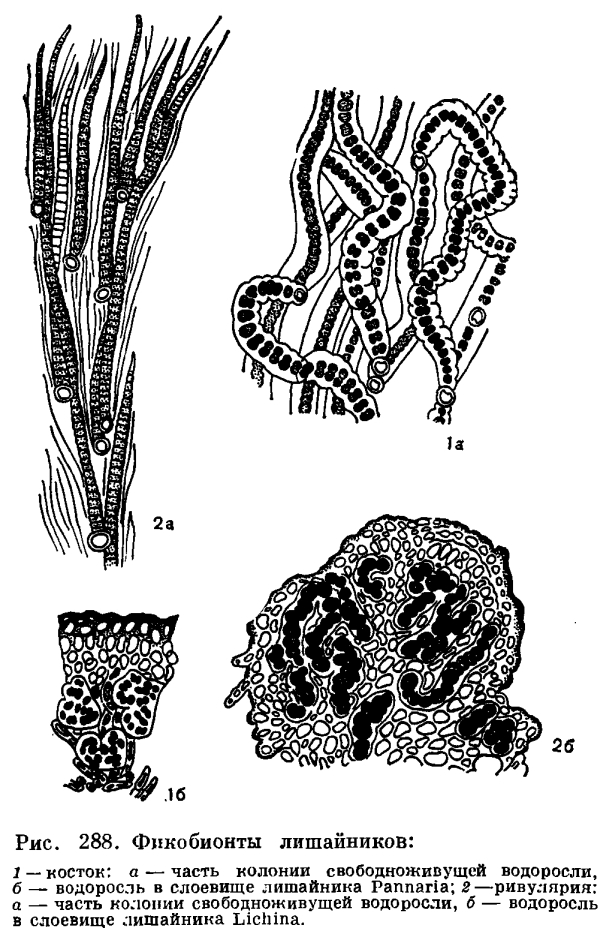
Водоросли в слоевище лишайника очень сильно изменяют свой внешний облик. Особенно это касается нитчатых водорослей, которые под влиянием гриба распадаются на отдельные клетки и часто деформируются до неузнаваемости (рис. 288). Размножаются водоросли внутри слоевища обычно делением и апланоспорами. Подвижных клеток размножения, характерных для свободноживущих водорослей, они не образуют. Но в лаборатории, в условиях культуры, фикобионты лишайников, как правило, принимают свой обычный, присущий им в свободноживущем состоянии облик и образуют подвижные зооспоры и гаметы.
Сине-зеленые водоросли как в слоевище лишайников, так и в лабораторных культурах образуют гетероцисты и гормогонии. Но число гетероцист в слоевище обычно бывает очень небольшим, гораздо меньшим, чем у свободноживущей водоросли.
Все вышеперечисленные особенности фикобионтов принципиально не отличают их от свободноживущих водорослей, при культивировании на искусственных средах имеющиеся отклонения быстро исчезают, и водоросли восстанавливают свой обычный внешний вид. По всей вероятности, эти изменения не закреплены генетически и возникают лишь под влиянием жизни водорослей в симбиозе.
Однако целый ряд физиологических особенностей, характерных для водорослей, живущих в слоевище лишайников, сохраняется у них и при культивировании на искусственных средах. Так, фикобионты лишайников отличаются замедлеппым ростом в слоевищах, эта же особенность сохраняется у них и при культивировании в лабораторных условиях. Причем если добавлять в культуры изолированных фикобионтов экстракты, полученные из микобионта лишайника, то рост водоросли усиливается. Интересно, что при таком медленном росте фотосинтетическая способность у лишайниковых водорослей ничуть пе меньше, чем у свободноживущих. В процессе своей жизнедеятельности они способны вырабатывать такое же количество углеводов, как и свободноживущие водоросли. Однако эти углеводы фикобионты используют не только для своего развития, значительное количество питательных веществ они отдают грибу (это и приводит к замедлению роста лишайниковых водорослей). Так, опытом было показано, что водоросль носток, выделенная из слоевища одного из видов коллемы и выращенная в культуре, выделяла в среду 5% азота, фиксированного из воздуха. Одновременно в среду поступали полисахариды и витамины (тиамин, биотин, рибофлавин, никотиповая и пантотеновая кислоты).
Было установлено, что водоросли в слоевище лишайников полностью или в значительной степени лишёны своих обычных избыточных ассимилированных ими веществ, крахмала и запасных веществ, липоидов, зерен цианофицина, гликогена и некоторых других.
Водоросли в слоевище лишайников способны переносить длительное высушивание. Опыты показали, например, что водоросль коккомикса в слоевище одного из лишайников выдерживала высушивание в течение 23 недель, а после нормализации условий восстанавливала все процессы своей жизнедеятельности. В культуре же этот фикобионт погибал без воды уже через 5 недель.
Все эти данные показывают, что в связи с особенностями жизни в контакте с грибом лишайниковые водоросли претерпевают не только морфологические, но и физиологические изменения.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Свидетельство и скидка на обучение каждому участнику
Занятие 2. Отдел лишайники – Phycomycota ( Lichenes)
Лишайники – симбиотические организмы, которые образованы грибом (гетеротрофный микобионт) и водорослями или цианобактериями (автотрофный фикобионт) (рис. 1). Гриб обеспечивает защиту водорослей от высыхания и действия крайних температур, а также снабжает их водой и минеральными солями. Водоросль снабжает гриб созданными ею органическими веществами. Такое сосуществование гриба и водоросли является постоянным, но не совсем равноправным – ведущую роль в симбиозе играет гриб. Гриб может питаться сапротрофно отмершими водорослями и продуктами их обмена, или как паразит, проникая внутрь клетки и поглощая ее содержимое (у низкоорганизованных лишайников). Поэтому партнерство в лишайнике является скорее не симбиозом, а контролируемым паразитизмом гриба на водоросли.


Рис. 1. Компоненты лишайника.
Грибной компонент каждого вида лишайника (в основном аскомицеты, но есть и базидиомицеты, фикомицеты) специфичен и отличается от микобионта других лишайников. Фотосинтезирующие организмы в составе лишайников представлены одноклеточными и нитчатыми зелеными водорослями (зеленые – требоксия, хлорелла, кладофора, плеврококк, трентеполия), а также цианобактериями (цистококк, глеокапса). В отличие от микобионта один и тот же вид водоросли может быть фикобионтом нескольких видов лишайников.
Вегетативное тело лишайников – слоевище, или таллом . Окраска слоевища обусловлена различными пигментами и специфическими органическими соединениями (лишайниковые кислоты) (рис. 2).


Рис. 2. Различная окраска таллома лишайников. Лишайниковые кислоты.
Последних около 300, и они откладываются на поверхности гиф в виде кристаллов, палочек, зернышек и придают талломам лишайников серую, сизую, зеленоватую, желтую, оранжевую, черную и др. окраску.
Основу вегетативного тела лишайников (плектенхиму) образуют переплетенные членистые гифы лишайниковых грибов. Некоторые гифы имеют толстую оболочку, способную разбухать, впитывая воду. Кроме того, имеются жировые гифы, содержащие капли жира.
По строение слоевища выделяют гетеромерные и гомеомерные лишайники

Рис. 3. Строение таллома гетеромерного лишайника: 1 – верхний коровой слой, 2 – клетка водоросли (цианобактерии), 3 – гонидиальный (альгальный) слой, 4 – сердцевинный слой, 5 – гиф гриба, 6 – нижний коровой слой, 7 – ризины (ризоиды).
У гомеомерных лишайников водоросли равномерно, по всей толще, распределены среди переплетающихся гиф грибов (рис. 4). Такое строение свойственно накипным и небольшой части листоватых и кустистых лишайников.

Рис. 4. Строение таллома гетеромерного лишайника.
По морфологии талломов различают три основные группы лишайников (рис. 5).

Накипные Листоватые Кустистые
Рис. 5. Морфологические группы лишайников.
Накипные, или корковые – тело в виде корочек или накипи, связанное с субстратом всей поверхностью и практически неотделимое от него. К этой группе принадлежит до 80 % всех лишайников . Выделяя лишайниковые кислоты, они разрушают поверхность скальные породы, подготавливая почву для заселения этой поверхности листоватыми и кустистыми лишайниками, мхами и затем цветковыми растениями. Некоторые из кислот имеют антибиотические или токсические свойства и, очевидно, выполняют защитную функцию.
Листоватые – тело в виде листовидных пластинок, прикрепленных к субстрату пучками гиф и легко отделяющихся от него. У слоевища можно отличить верхнюю и нижнюю поверхности. Они являются пионерами растительных сообществ (пармелия, ксантория, центрария).
Кустистые – таллом в виде более или менее разветвленного кустика, поднимающегося с земли, или свисающего с ветвей (ягель, или олений мох, уснея, кладония, рамалия) .
Размножаются лишайники преимущественно вегетативно – обломками слоевища или специальными органами – соредиями и изидиями (рис. 6) . Отделение участков слоевища происходит механически (особенно в сухую погоду, когда они становятся хрупкими).


Рис. 6. Вегетативное размножение лишайников. 1 – соредии, 2 – изидии, 3 – выброс соредиев.
Соредии образуются внутри водорослевого слоя и состоят из небольшого числа клеток водоросли и оплетающих их гиф гриба. Через разрыв в верхней коре слоевища соредии выпадают наружу и разносятся ветром, в благоприятных условиях они образуют новый таллом.
При половом размножении на слоевищах лишайников в результате полового процесса формируются половые спороношения в виде плодовых тел. Большинство лишайников формируют открытые плодовые тела в виде апотециев – дисковидных образований (рис. 7). Обычно апотеции образуются на верхней поверхности слоевища лишайников, плотно прирастая к ней своей нижней частью или возвышаясь над ней на тонкой маленькой или довольно длинной ножке. У некоторых лишайников они бывают целиком погружены в слоевище.





Рис. 7. Плодовые тела лишайников: апотеции (1), перитеции (2), гастеротеции (3): морфология и строение.

Апотеции разделяют на леканоровые (довольно мягкие, с развитым слоевищным краем), лецидеевые (относительно твердые, с собственным краем) и биаторовые – переходный тип между леканоровыми и лецидеевыми (рис. 8).
Рис. 8. Типы апотеций. 1 – лецидеевые, 2 – биаторовые, 3 – леканоровые.
Перитеции – закрытые плодовые тела, имеющее вид маленького кувшина с отверстием наверху.
Небольшое количество лишайников образуют узкие плодовые тела удлиненной формы, которые называют гастеротециями. Обычно гастеротеции имеют вид штрихов, простых или разветвленных линий.
В апотециях, перитециях и гастеротециях споры развиваются внутри сумок – особых мешковидных образований. Лишайники, формирующие споры в сумках, объединяются в большую группу сумчатых лишайников. Они произошли от грибов класса аскомицетов. У небольшой группы лишайников споры образуются не внутри сумок, а экзогенно, на вершине удлиненно-булавовидных гиф – базидий. Лишайники с таким образованием спор объединяются в группу базидиальных лишайников. При прорастании одноклеточной споры образуется одна, редко до пяти ростковых трубочек. Ростковые трубочки растут, разветвляются и образуют первичный мицелий, который еще не содержит водорослей. Дальнейшее развитие этого мицелия и образование слоевища лишайника происходит лишь в том случае, если его гифы встречают водоросль, соответствующую данному виду лишайника.
Бесполые спороношения. Помимо спор, образующихся половым путем, у лишайников известны также бесполые спороношения – конидии, пикноконидии и стило-споры, возникающие экзогенно на поверхности конидиеносцев (рис. 9). При этом конидии образуются на конидиеносцах, развивающихся непосредственно на поверхности слоевища, а пикноконидии и стило-споры в особых вместилищах – пикнидиях.

Рис. 9. Органы бесполого спороношения (1- конидиеносцы, 2 и 3 - пикнидии)
Рост лишайников происходит очень медленно. За год их таллом нарастает у разных видов от 1 до 10 мм. Продолжительность жизни таллома у отдельных видов может достигать 100 лет и более.
По отношению к субстрату и факторам окружающей среды лишайники подразделяют на ряд экологических групп: эпигейные – растут на почве, эпифитные – на живых растениях, эпиксильные – на обработанной или гниющей древесине, эпилитные – на камнях. Основное условие поселения лишайников – длительная неподвижность субстрата.
В лишайниках накапливаются полисахариды, а белков и жиров мало. Некоторые химические вещества лишайников обладают антимикробными свойствами. Кроме того, представители этого отдела используются для получения антибиотиков (кладония, пармелия, эверния и др.), ароматических веществ (некоторые лишайники синтезируют ценное эфирное масло) и фиксаторов запахов (лобария, эверния), а также красителей шерсти.
Большинство лишайников не выдерживают даже малейшего загрязнения воздуха, их можно использовать для общей оценки степени загрязненности окружающей среды. На этом основано одно из направлений индикационной экологии – лихеноиндикация.
Водоросли, встречающиеся в слоевище лишайников, называются фикобионтами лишайников. По своему систематическому отношению они относятся к различным отделам: к сине - зеленым, зеленым, желто- зеленым и бурым водорослям.
Как показало изучение лишайниковых фикобионтов, в слоевище лишайников встречаются представители 28 родов водорослей.
Водоросли в слоевище лишайников очень сильно изменяют свой внешний облик. Размножаются водоросли внутри слоевища обычно делением или апланоспорами.
Лишайниковые водоросли очень устойчивы к высоким температурам, способны переносить длительное высушивание.
Взаимоотношения гриба и водоросли в теле лишайника
Вопрос взаимоотношения гриба и водоросли в слоевище лишайника с давних пор волнует лихенологов. Со дня открытия С. Швендера прошло более 100 лет. За этот период появилось не менее десятка теорий, пытающихся объяснить отношение между грибом и водорослью, однако среди них нет ни одной общепризнанной и окончательно доказанной. С.Швендер, обнаружив, что лишайник состоит из гриба и водоросли, предположил, что гриб в слоевище паразитирует на водоросли. Однако он ошибочно отвел роль хозяина грибу, а водоросли - раба.
Но уже в те времена некоторые ученые выдвинули мысль о двустороннем паразитизме компонентов лишайника – гриба на водоросли и водоросли на грибе. При этом было высказано предположение, что гриб и водоросль в слоевище лишайника находятся в полном морфофизиологическом единствеи связаны между собой так же, как корни и листья цветковых растений.
Однако в 1873г. этой идеалистической теории был нанесен удар. Известный французский исследователь Е. Борне обнаружил внутри водорослевых клеток грибные отростки – гаустории, всасывающие органы гриба. Это позволяло думать, что гриб использует содержимое клеток водорослей, т. е. ведет себя как паразит.
Установили, что гриб получает вещества, ассимилируемые водорослью. Однако для существования как самого гриба, так и лишайника в целом необходимо, чтобы водоросль, окруженная со всех сторон грибными гифами, все-таки могла жить и более или менее нормально развиваться. Если гриб начнет проявлять себя слишком активно, поражать все без исключения водоросли, то погибнет и сам, а значит, перестанет существовать и лишайник.
Гриб должен использовать лишь часть водорослей, оставляя резерв – здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Способность водорослей нормально развиваться и даже размножаться в слоевище лишайника сохраняется благодаря умеренности паразитизма самого гриба.
Отношения между мико- и фикобионтом в слоевище лишайника не сводятся только к паразитизму гриба на водоросли. Еще в начале нашего века крупнейший русский лихенолог А.А.Еленкин обнаружил в их слоевище некральные зоны водорослей – скопления отмерших, потерявших зеленую окраску клеток, расположенные несколько ниже зоны живых водорослей. К этим бесцветным мертвым клеткам водорослей тоже тянулись грибные гифы. Это привело Л. А. Еленкина к мысли, что гриб, убив водоросль, переходит к сапрофитному способу питания, поглощая и ее мертвые остатки. Таким образом, гриб в слоевище лишайника ведет себя и как паразит, и как сапрофит. И отношения между грибом и водорослью в слоевище лишайника Л. А. Еленкин назвал эндопаразитосапрофитизмом.
В 60-х годах нашего столетия крупнейший советский лихенолог А.Н.Окснер пришел к выводу, что водоросль в слоевище лишайника обязательно должна забирать у грибного компонента все необходимые для своего существованиявещества, за исключением тех органических соединений, которые она сама вырабатывает на свету. К этим веществам относится вода и минеральные соли, азотистые и некоторые другие неорганические соединения. Следовательно, и водоросль в слоевище лишайника проявляет себя как паразит. Причем это вовсе не противоречит общему характеру ее питания. Многие из водорослей способны и к миксотрофному питанию.
Таким образом, ученые считают, что водорослевый и грибной компоненты лишайника находятся в очень сложных взаимоотношениях. Микобионт ведет себя как паразит и сапрофит на теле водоросли, а фикобионт паразитирует на лишайниковом грибе. Однако все высказанное по этому поводу до сих пор остаётся лишь догадкой. Ученые пока не научились выращивать и поддерживать в живом состоянии слоевище лишайников в искусственных условиях.
Одна из причин неудач подобных попыток - чрезвычайно медленный рост лишайников. Лишайники – многолетние растения. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20 – 50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2 – 0,3мм.
Кустистые и листоватые лишайники растут несколько быстрее – вгод их слоевище увеличивается на 2 – 3мм. Поэтому, чтобы вырастить взрослый лишайник в лаборатории, требуется не менее 20 лет. Трудно проводить столь долговременный эксперимент!
Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов лишайников, остаются пока лишь догадками.
Читайте также: