
Мегаспорангием у покрытосеменных растений является
Обновлено: 04.07.2024
В мегаспорангии эфедры из мегаспоры в результате свободного ядерного деления и последующего образования клеточных перегородок развивается массивный женский гамето-фит. В каждом из них закладываются обычно два архегония. Архегоний имеет длинную шейку, состоящую из 32 или более клеток. При делении ядра центральной клетки архегония между брюшным канальцевым ядром и ядром яйцеклетки не образуется клеточной перегородки.[ . ]
У примитивных групп голосеменных мегаспорангий (т. е. нуцеллус семязачатка) открывается посредством сквозных пыльцевых камер, Мужские гаметы у таких голосеменных представляют собой подвижные сперматозоиды, которые с помощью многочисленных жгутиков легко достигают женского гаметофита и проникают в архегоний.[ . ]
У некоторых селагинелл не найдены микро-или мегаспорангии. У других, по-видимому, апогамных видов спорангии содержат не четыре одинаковые, как свойственно большинству видов, а одну нормальную и три абортивные мегаспоры.[ . ]
Микроспоры образуются в микроспорангиях, а мегаспоры — в мегаспорангиях (нуцеллусах) семязачатков. Микроспорангии развиваются на микроспорофиллах, а семязачатки — на мегаспорофиллах. Микро- и мегаспорофиллы голосеменных имеют различный внешний вид, размеры и строение. В одних случаях они крупные и перистые, в других более мелкие и цельные. У хвойных далеко зашедшие процессы редукции и срастания привели к тому, что мегаспорофиллы стали практически неразличимы в общей слитной структуре так называемой семенной чешуи.[ . ]
Женский гаметофит у всех голосеменных развивается полностью внутри мегаспорангия и не выходит даже частично наружу, т. е. не соприкасается непосредственно с воздушной средой. Доступ к женскому гаметофиту осуществляется только через микрониле. Таким образом внутри семязачатка создаются наиболее благоприятные условия защиты женского гаметофита от высыхания. В результате происходит постепенная редукция и упрощение женского гаметофита и архегониев, возникает возможность очень раннего формирования яйцеклетки, и у некоторых голосеменных (вельвичия и гнетум) образуются даже особые нео-тенические безархегониальные гаметофиты.[ . ]
Это и есть собственно мегаспорангий, который, в отличие от открытых микроспо-рангиев саговниковых, как и у других голосеменных, заключен в защитный покров.[ . ]
У некоторых паразитных двудольных покров семязачатка исчезает полностью и мегаспорангий оказывается, таким образом, голым. Такие а т е г м а л ь и ы о (от греч. а •— частица отрицания) семязачатки характерны для многих, представителей порядка санталовых. У некоторых баланофоровых утрачен не только покров семязачатка, но даже стенка мегаспорам гия.[ . ]
Мужские и женские гаметофиты развиваются из микро- и мегаспор, которые образуются в микро- и мегаспорангиях, заключенных в микро- или мегасорусах. Сорусы образуются па спороносных сегментах погруженного листа. Мегасорусы, обычно на ножках, закладываются первыми у основания сегмента. Микросорусы, как правило, сидячие и размещаются на остальной части сегмента.[ . ]
Споры развиваются в микро- и мегаспорангиях, собранных в сорусы и заключенных в замкнутые вместилища, которые получили название спорокарпиев (рис. 148).[ . ]
У разносноровых высших растений (развивающих гетероспоры) число микроспор в каждом микроспорангии также обычно не бывает меньше 32, но в мегаспорангии, как правило, образуется только одна мегаспора.[ . ]
Каждый сорус окружен нежным покрываль-цем, и два ряда сорусов заполняют всю полость спорокарпия. Ложе соруса гребневидное и несет сверху ряд мегаспорангиев, а по бокам многочисленные микроспорангии. Заложение спорангиев идет от верхушки ложа к основанию (сорусы градатного типа). У некоторых видов встречаются сорусы смешанного типа. Число сорусов в спорокарпии варьирует от двух у марсилеи египетской до 12 у марсилей покрытой и четырехлистной. Соотношение микроспорангиев и мегаснораигиев внутри одного соруса — также непостоянная величина. Встречаются экземпляры марсилей, в сорусах которых полностью отсутствуют мегаспорангии (особенно часто у марсилей маленькой и покрытой). Как правило, в микроспорангиях развивается по 64 споры, в то время как в мегаспорангиях — по одной крупной мегаспоре. Спорангии снабжены ножкой и однослойной стенкой. Рядом признаков они напоминают спорангии схизейных.[ . ]
Микроспорангий саговниковых по внешнему виду и внутреннему строению значительно меньше отличается от микроспоравгиев папоротниковидных, чем это проявляется у мегаспорангиев.[ . ]
Образование семени является первым этапом в развитии спорофита. Женские шишки построены из крупных чешуек, называемых мегаспорофиллами, каждая из которых несет по .два мегаспорангив на внутренней поверхности, а каждый мегаспорангий в свою очередь содержит мегаспору, которая развивается в многоклеточный гаметофит, содержащий две или три архегонии. Каждая архегония состоит из одиночной большой яйцеклетки и нескольких маль1х вытянутых клеток. Мегаспорангий покрыт так называемым интегу-ментом. Мегаспорангий с интегументом называют семязачатком.[ . ]
Почковидные или обратнояйцевидные спорангии сидят на ножках на оси стробила близ пазухи или в пазухе спорофилла; с той стороны ножки, что обращена к верхушке спорофилла, располагается язычок. Мегаспорангии обычно много крупнее микроспорангиев (рис. 62).[ . ]
Среди современных голосеменных пыльцевые камеры хорошо выражены у саговниковых и у гинкго. У более высокоспециализированных голосеменных, таких, как хвойные, эфедра (хвойник), вельвичия и гнетум, мегаспорангии не раскрываются и пыльцевая трубка, несущая неподвижные мужские гаметы (спермии), активно прободает их стенку на своем пути к архегонию.[ . ]
Эволюция папоротникообразных привела к появлению и развитию так называемой разноспоровости. Споры и производящие их спорангии становились разными. Появились микроспорангии, мелкие споры которых прорастают в мужской гаметофит, и мегаспорангии, споры которых прорастают в женский гаметофит. У всех папоротников, плаунов и хвощей нет главного корня, а есть только придаточные корни на корневищах.[ . ]
Стробилы лепидодендронов, сидевшие на концам ветвей, могли достигать в длину 50 см и диаметра 5 см. В обоеполых стробилах мегаспорофиллы располагались в нижней, а микроспорофиллы — в верхней их части. В микроспорангиях развивались тысячи мелких микроспор. У более примитивных видов лепидодендронов в мегаспорангиях развивались сотни небольших мегаспор. У более подвинутых их число уменьшилось до 16, даже до 12 и 4, а размер мегаспор увеличился почти до 1 мм. И наконец, в спорангиях наиболее подвинутых видов развивалась всего одна мегаспора, которая тут же прорастала, образуя женский га-метофит (заросток), защищенный от неблагоприятных воздействий внешней среды толстой стенкой спорангия или завернутыми краями спорофилла, создававшими для гаметофита дополнительный покров (рис. 58).[ . ]
Сальвиния ушковидная (За№п1а аиг!сиЫа): 1 — мутовка листьев: а — плавающие листья, б — сегменты погруженного листа, в — сорусы; г — плавающий лист; з — сосочек листа с 4 волосками. Сальвииия продолговатолистиан (8. оЫощпГоНа): Ю — надводный лист; и — плавающий лист в поперечном разрезе: а — листовая пластинка, б — киль.[ . ]
Эти два тина не всегда бывают строго разграничены, и между ними известны различные переходы. Первый тип является более примитивным, второй тип возник из первого в результате редукции мега-спорангия.[ . ]
Образовавшаяся в нуцеллусе в результате мейоза единственная функционирующая мегаспора быстро растет за счет трех остальных, вскоре отмирающих спор тетрады и окружающих клеток нуцеллуса и вырабатывает хорошо выраженную двуслойную оболочку, как это свойственно спорам бессемейных высших растений, переносимым токами воздуха. Но мегаспора саговниковых никогда не покидает мегаспорангия, и назвапная особенность ее строения сохранилась как пережиток прошлого, унаследована от далеких предков, расселявшихся с помощью спор. У саговников наружная оболочка мегаспоры в дополнение пропитывается кутином, что, очевидно, придает ей еще более архаичный характер.[ . ]
Когда пыльца попадает на женские шишки, она проходит в семязачаток, причем каждая пыльцевая гранула развивается в тычиночную трубочку и два спермоядра, а когда тычиночная трубочка проникает в яйцеклетку, происходит слияние спермоядра с ядром яйцеклетки. Это и есть оплодотворение. Диплоидная зигота становится диплоидным зародышем. Со временем внешний инте-гумент семязачатка превращается в оболочку семени, а из остатков мегаспорангия образуется эндосперм. После созревания семена из шишек выпадают наружу.[ . ]
В микросорусах сальвинии ушковидной образуется до 500 микроспорангиев. В каждом микроспорангии образуется 32 (у сальвинии плавающей — 64) микроспоры. Микроспоры имеют ясно выраженный трехлучевой рубец. Созревшие микроспоры по 4 погружены в отвердевшую плазматическую массу, называемую массулой. Замечателен тот факт, что ископаемая сальвиния Рёйза (Б. геийи) обладала амфиспорангиатными сорусами с многочисленными микроспорангиями и меньшим числом мегаспорангиев. У современных сальвиний были отмечены случаи развития наряду с нормальными не свойственных им амфиспорангиатных сорусов.[ . ]
Однако большинство ботаников справедливо считают, что наличие у цветковых мегаспор ангин, правда очень упрощенного и редуцированного, не вызывает сомнений. Ведь в нуцеллусе имеются два необходимых элемента каждого спорангия — спорогепная ткань (обычно крайне редуцированная) и стенка, которая, однако, гистологически сильно упрощена. Совершенно очевидно, что нуцеллус цветковых морфологически вполне соответствует нуцеллусу голосеменных, т. е. они представляют собой гомологичные образования и являются мегаспорангиями.[ . ]
Класс 6—хвойные, или пннопсиды (Pinopsida). Наиболее многочисленная группа среди современных голосеменных, геологическая история которой восходит к раннему карбону. Листья у современных форм цельные, с одной жилкой или со слабо развитым дихотомическим жилкованием, но среди вымерших форм известны вильчатые листья. Редуцированные спорофиллы собраны в однополые стробилы. Мужские гаметы лишены жгутиков. Делится на два подкласса — вымерший подкласс кордаитиды (Cordaitidae) и современный подкласс хвойные (Pinidae).[ . ]
В верхнедевонском периоде истории Земли возникли и распространились достигшие максимального развития в каменноугольном периоде семенные папоротники. Они известны только по ископаемым остаткам. Эти растения явились как бы новаторами в эволюции наземной флоры. В их внешнем облике много общего с папоротниками, и прежде всего крупные листья с многократно рассеченными листовыми пластинками. Но они существенно отличались от папоротников тем, что на их листьях формировались зачатки семян — семяпочки. По общему плану строения эти органы вполне соответствовали семяпочкам современных голосеменных. Здесь был покров (интегумент) с отверстием наверху (микропиле); в большей своей части он обычно срастался с наружной поверхностью покрываемого им массивного мегаспорангия, оставаясь свободным только наверху. В мегаспорангии, который у голосеменных обычно называют ну-целлусом, развивалась только одна мегаспора, которая здесь же и прорастала. В результате этого внутри мегаспоры образовывался мелкоклеточный заросток. Однажды в нем были найдены окаменевшие архегонии.[ . ]
При прорастании микроспоры содержимое ее делится на маленькую проталлиальную клетку и большую антеридиальную клетку. Ядро антеридиальной клетки делится на 2 ядра, одно из которых представляет собой ядро трубки, а второе — ядро генеративной клетки. Вокруг ядра генеративной клетки имеется слой цитоплазмы, и мы можем поэтому говорить о генеративной клетке. Ядро трубки первым входит в пыльцевую трубку, а за ним следует генеративная клетка. Таким образом, зрелое пыльцевое зерно состоит из проталлиальной клетки, генеративной клетки и ядра трубки. При прорастании пыльцевого зерна генеративная клетка входит вслед за ядром трубки в пыльцевую трубку и делится при этом на две неравные мужские клетки (или ядра), меньшая из которых, по-видимому, дегенерирует. Микроспоры переносятся, вероятно, насекомыми на капельку жидкости, выделяемой микропиляр-ной трубкой. Здесь они втягиваются высыхающей жидкостью до мегаспорангия, причем пыльцевая трубка образуется часто еще в микропи-лярной трубке.[ . ]
В семязачатках наблюдаются два типа строения мегаспорангия и в соответствии с этим различают два типа семязачатков, названные Ф. Ван Тигемом (1898) крассинуцеллятными (от лат. crassus — толстый и nucellus — ядрышко) и тенуинуцеллятными (от лат. tennis — топкий). Крассинуцеллятные семязачатки характеризуются относительно мощно развитым мегаспорангием, и поэтому материнская клетка мегаспор (мегаспороцит) отделена от эпидермы мегаспорангия одним или несколькими слоями клеток. У тенуинуцеллятных семязачатков, мегаспорангий которых более редуцирован, материнская клетка мегаспор лежт непосредственно под эпидермой мегаспорангия. Эти два типа не всегда бывают строго разграничены, и между ними известны различные переходы. Первый тип является более примитивным, второй тип возник из первого в результате редукции мегаспорангия. Поэтому крассинуцеллятные семязачатки характерны для относительно более примитивных двудольных и для большинства однодольных, в то время как тенуинуцеллятные семязачатки встречаются у относительно более подвинутых двудольных и лишь у немногих однодольных. Кроме того, крассинуцеллятные семязачатки чаще всего бывают битегмальными, а тенуииуцоллятиые, как правило, унитегмальные.
У цветковых растений в мегаспорангии образуется обычно одна материнская клетка мегаспор. Она претерпевает два деления мейоза, в результате чего аналогично появлению тетрады гаплоидных микроспор образуется тетрада гаплоидных мегаспор. Как и при формировании микроспор, появляются каллозовые оболочки, обеспечивающие изоляцию материнской клетки и мегаспор от окружающих клеток.
Мегаспоры тетрады бывают расположены поразному, но чаще всего образуется линейная тетрада, реже — Т-образиая и еще реже — квадратная или тетраэдрическая. У громадного большинства цветковых растений возникают четыре мегаспоры, из которых нижняя (халазальная) и значительно реже верхняя (микропилярная), или промежуточная, увеличиваются в размерах и функционируют как мегаспора, а остальные три отмирают и разрушаются. Очень редко вместо четырех потенциальных мегаспор образуются три, две, а иногда только одна. В последнем случае материнская клетка мегаспоры становится непосредственно мегаспорой.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру: от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.

Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) - важнейшими производителями органических веществ на суше, как водоросли - в морях и океанах.
Цветок - генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения. Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище - завязи, сформированной из плодолистика (-ов).
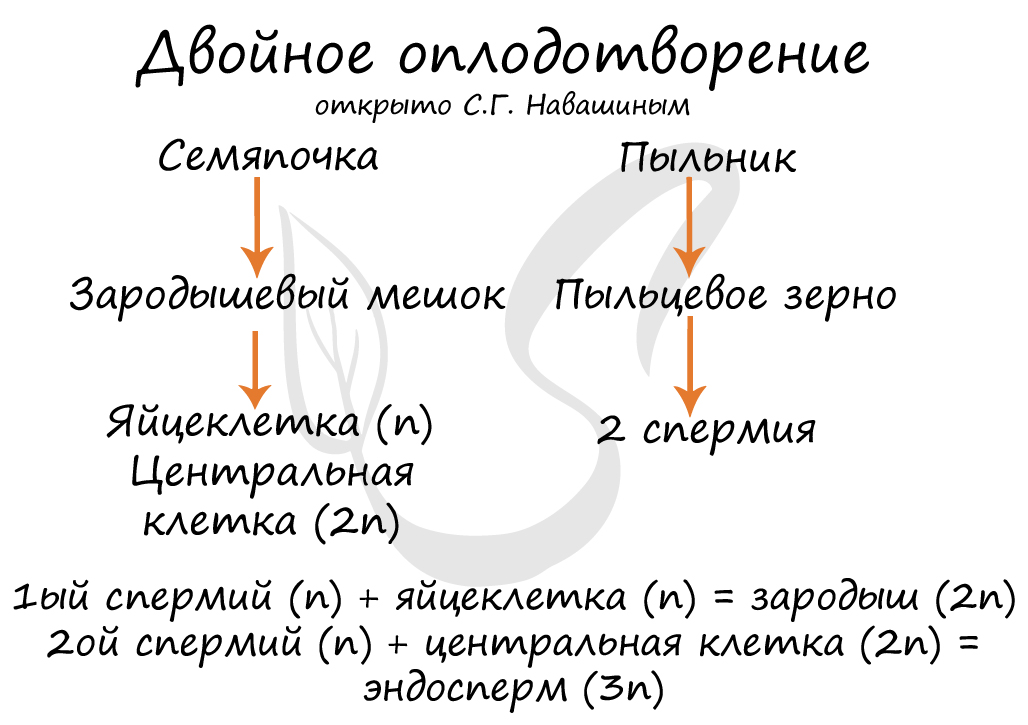
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества - эндосперма (3n). Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод - генеративный орган, служащий для защиты и распространения семян.
Ксилема - проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами, а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

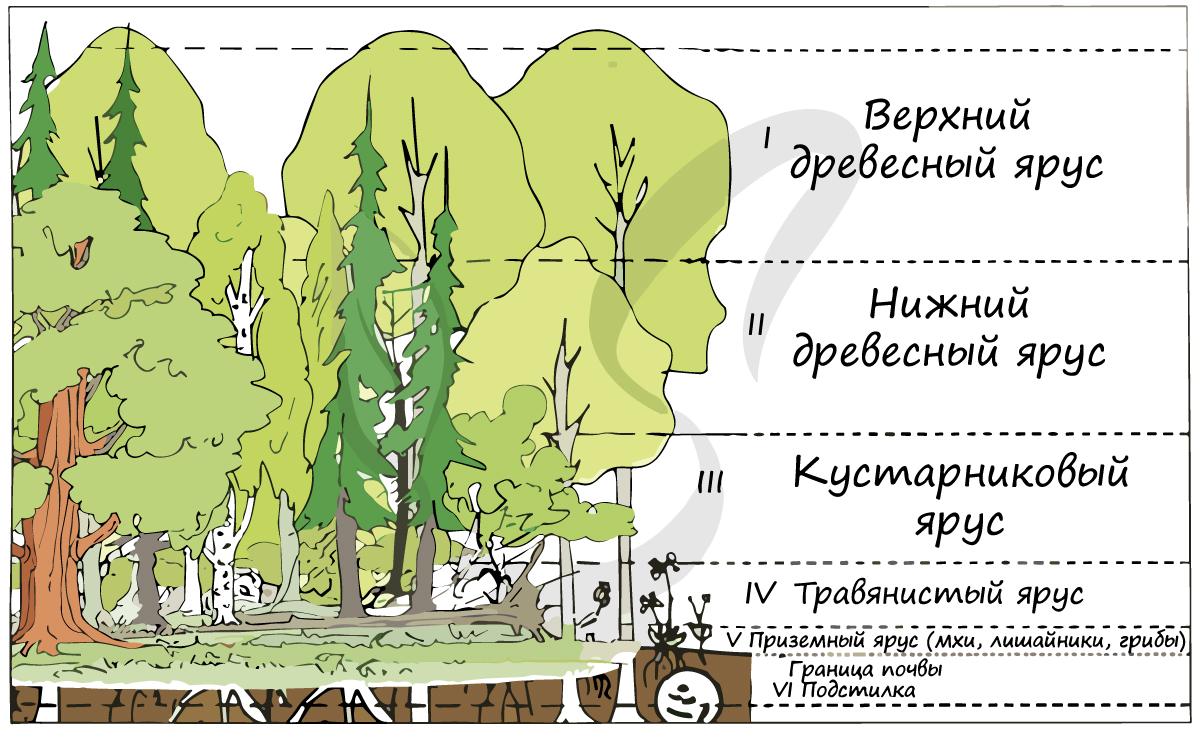
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых :)
Классы покрытосеменных
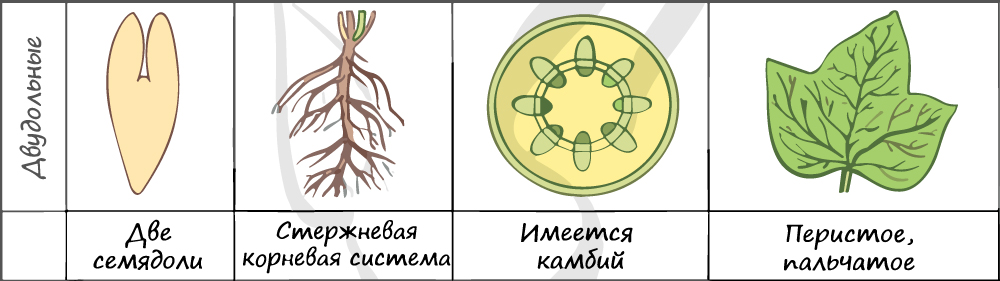
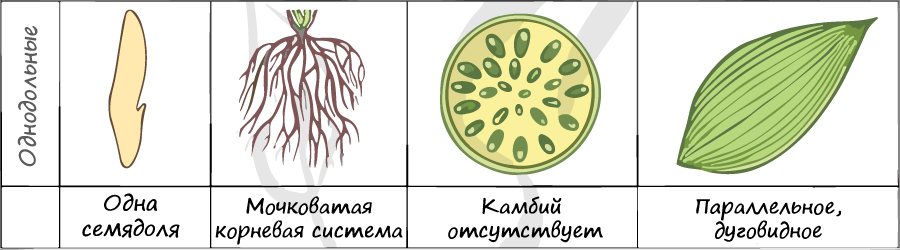
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные, бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
-
Двудольные - семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
-
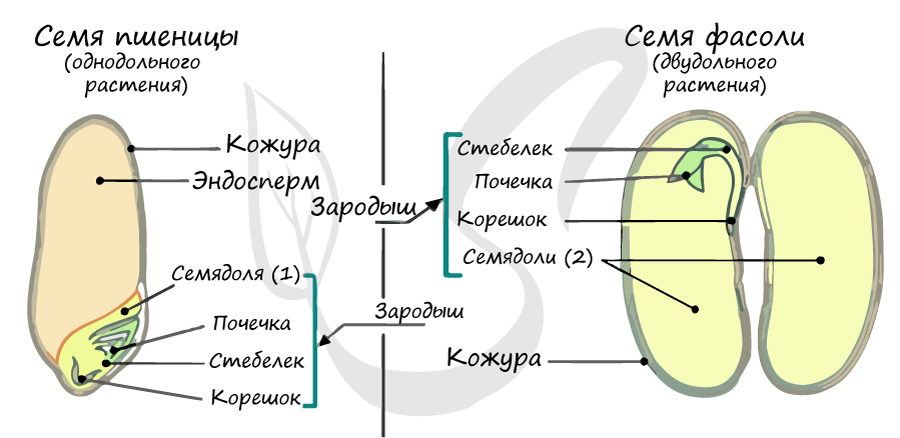
В составе зародыша обычно имеется две семядоли
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни. Главный корень развивается из зародышевого корешка.
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

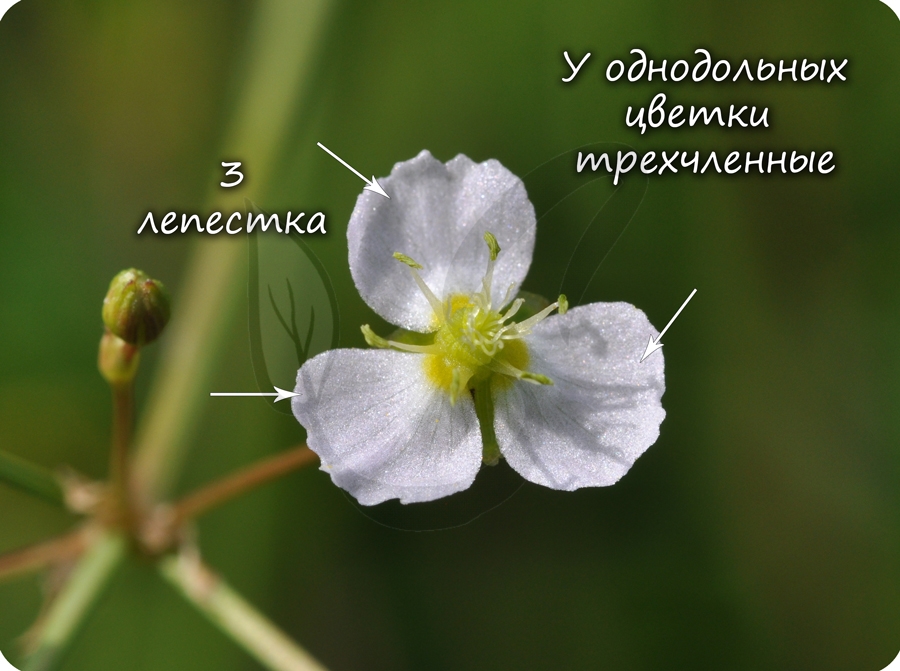
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
Эндосперм семени
Эндосперм (от греч. endon - внутри + греч. sperma - семя) - запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у - березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
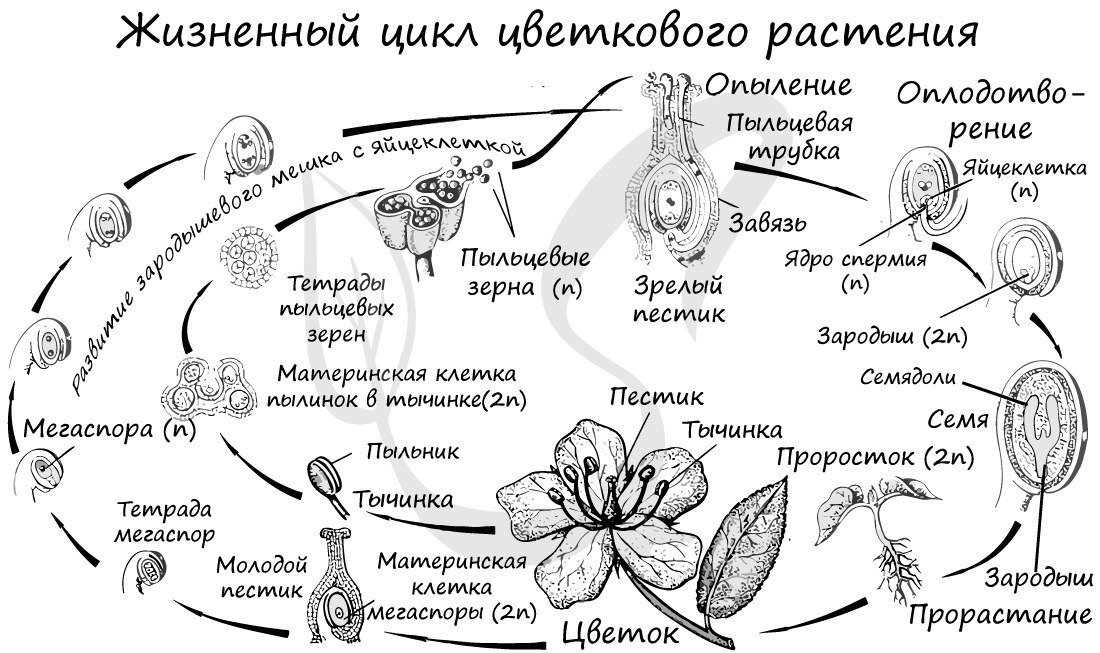
Жизненный цикл
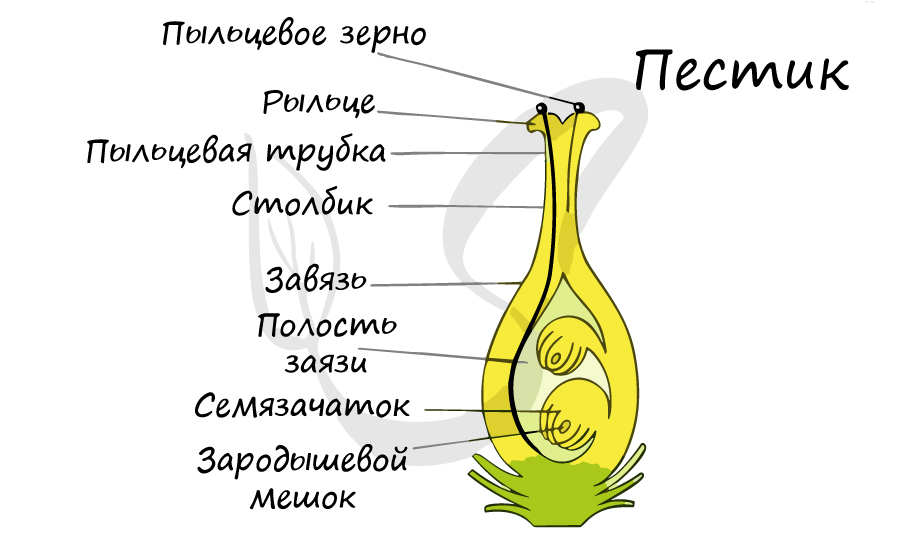
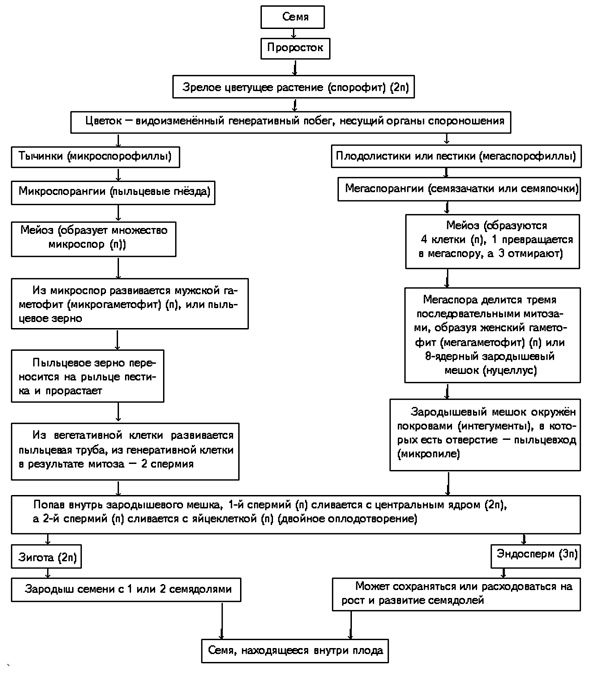
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n) в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) - мужской гаметофит. В завязи пестика в семязачатке формируется женский гаметофит - зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток. Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) - запасного питательного вещества. Другой спермий (n) сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник - образуется плод. Своим внешним видом плоды привлекают животных, и те их охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия, они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте - почти все культурные растения принадлежат к этому отделу! Цветковые имеют медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Итак, в прошлый раз мы учили много умных слов: микроспора, женский гаметофит, антеридий и т.д. И если по отношению к споровому растению все эти слова имеют однозначное наименование, то с отделом Покрытосеменных все немного по-другому. У его представителей каждая из этих структур имеет и традиционное название.
Покрытосеменные - разноспоровые растения, поэтому вспомним схему жизненного цикла разноспорового плауна селагинеллы:
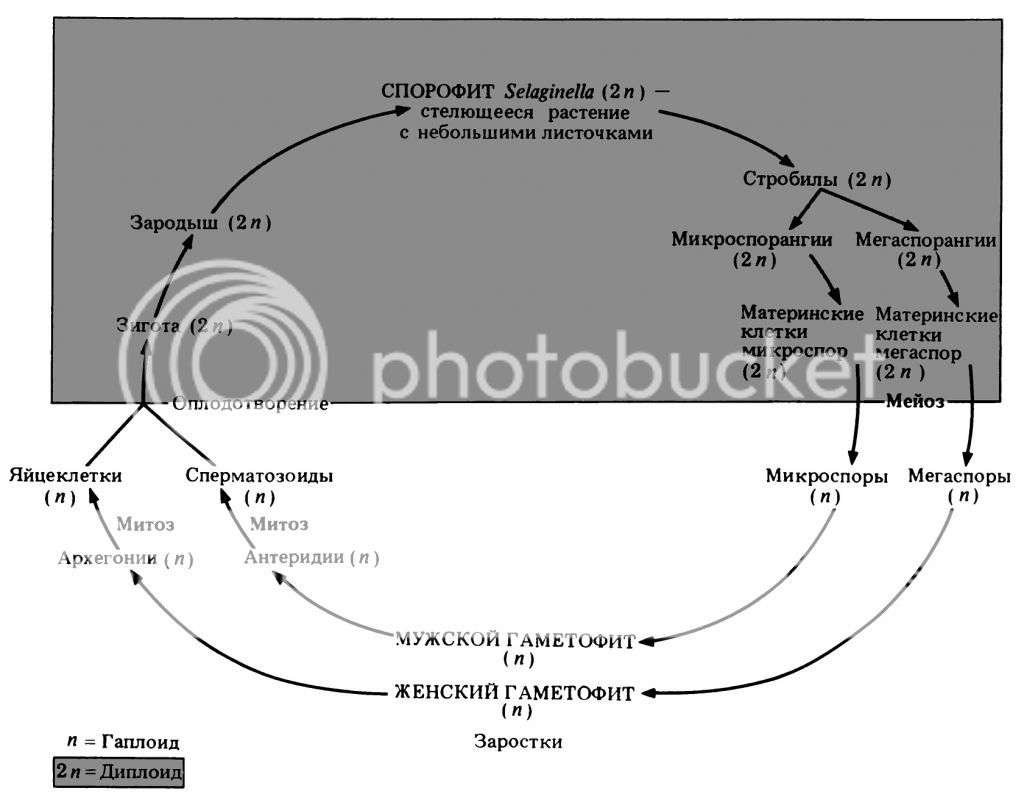
Такой же жизненный цикл будут иметь Покрытосеменные растения. Теперь осталось подобрать для каждой стадии соответствующую структуру Покрытосеменного растения.
Спорофит - и так понятно, взрослое растение. Далее нужно найти спорангии, сидящие на стробилах. Стробилы - это генеративные побеги, на которых имеются спорофиллы - листья, несущие спорангии, иными словами - спороносные колоски. Генеративный побег у нас уже есть - это цветок. За генеративную функцию у него отвечают пестики и тычинки. Вот это и есть наши спорофиллы.
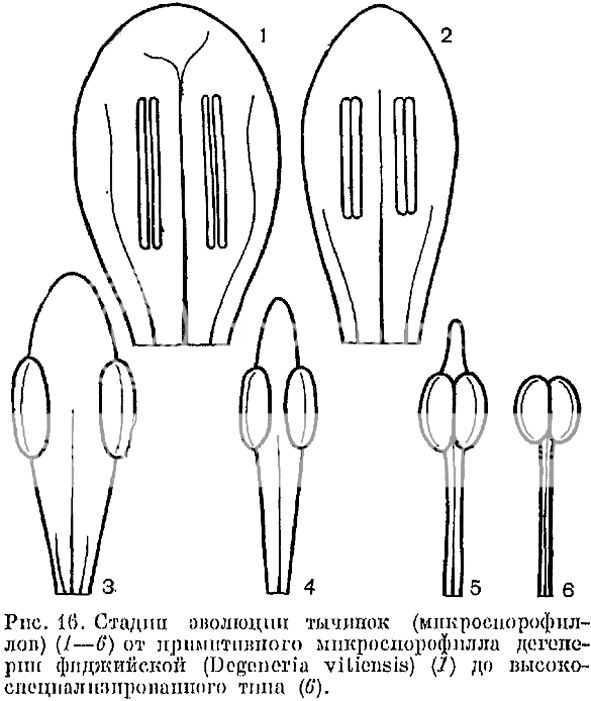
Эволюция тычинок: сначала широкое основание листа, несущего, как и у спорового растения, спорангии, затем листовая пластинка сужается, количество жилок уменьшается, постепенно остается практически одна лишь центральная жилка, на которой сидят спорангии.

Эволюция пестиков: вначале были плодолистики. Это листики, несущие спорангии, и при этом свернувшиеся с местом перегиба по средней жилке, так что спорангии оказались внутри этого свертка. Эту стадию мы и видим на рисунке 1. Затем мы уже можем преобразовывать щель между двумя сторонами спорофилла, которая постепенно замыкается и становится улавливающей поверхностью рыльца.
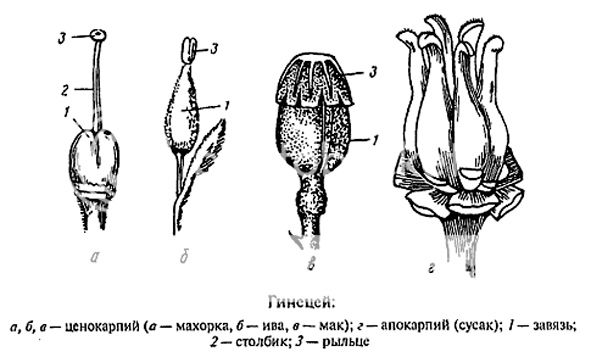
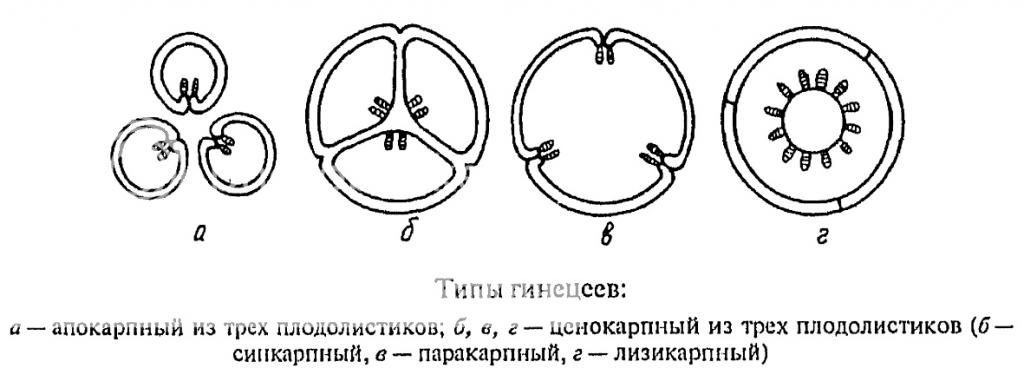
Совокупность плодолистиков мы называем гинецей. Гинецей может состоять как из отдельных плодолистиков, как на рисунке г), так плодолистики могут срастаться и образовывать пестики с единой (внешне, только внешне) завязью, при дальнейшем срастании может образовываться единый столбик и рыльце. Чаще всего количество лопастей на рыльце может указывать на количество плодолистиков в пестике. Различные типы срастания плодолистиков можно посмотреть на следующем рисунке.
Так как плодолистик это мегаспорофилл (лист, на котором образуются мегаспорангии), то теперь можно на нем найти мегаспорангий. Семяпочка? Не совсем. Мегаспорангий у Покрытосеменных покрыт двумя оболочками, которые называются интегументы. Сам мегаспорангий называется нуцеллус.
Все, путь от мегаспорангия до яйцеклетки пройден.
Теперь разберемся с микроспорангиями.
Мы уже выяснили, что тычинка у нас это микроспорофилл, т.е. лист, несущий микроспорангии. На каждом листе имеется два пыльника, каждый из который состоит из двух пыльцевых мешков. Вот пыльцевой мешок - это и есть микроспорангий. В нем образуется спорогенная ткань (т.е. множество материнских клеток спор), которая делится путем мейоза и образует гаплоидные микроспоры.
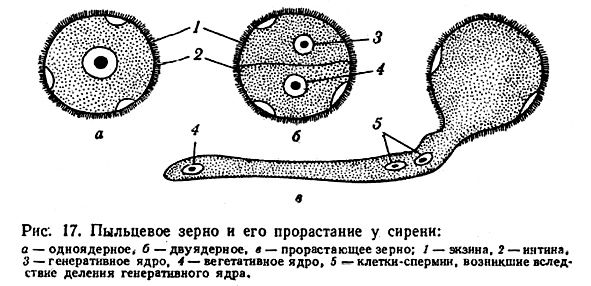
Микроспоры - это и есть пыльцевое зерно (рисунок а). Но здесь тут же начинает развиваться мужской гаметофит, сначала состоящий из 2 клеток (рисунок б), а потом из 3 (рисунок в). Опять же, речи о возникновении антеридиев не идет, т.к. гаметофит слишком мал. При опылении пыльцевое зерно попадает на рыльце пестика, в это время оно может представлять собой любую стадию от микроспоры до 3-клеточного гаметофита. Клетка пыльцевой трубки прорастает, два спермия (1n) достигают зародышевого мешка и происходит двойное оплодотворение, которое является специфическим для покрытосеменных растений. При оплодотворении яйцеклетки (1n) образуется зигота (1n + 1n = 2n), при оплодотворении центральной клетки (2n) образуется первичная клетка эндосперма (1n + 2n = 3n). Из зиготы образуется зародыш семени, из первичной клетки эндосперма - соответственно, эндосперм.
Что из всего этого нужно знать? Пожалуй, двойное оплодотворение вместе с плоидностью клеток. Все остальное - это уже для олимпиад и для дополнительных вступительных испытаний в престижные ВУЗы.
Читайте также: