
Проводящая система представлена сосудами у голосеменных или покрытосеменных
Обновлено: 05.10.2024
Свидетельство и скидка на обучение каждому участнику
Отдел Голосеменные
Общая характеристика. Первые голосеменные появились в конце девонского периода около 350 млн. лет назад; вероятно, они произошли от древних папоротниковидных, вымерших в начале каменноугольного периода. Отдел современных голосеменных насчитывает более 700 видов. Несмотря на относительно малую численность видов, голосеменные завоевали почти весь земной шар. В умеренных широтах Северного полушария они на огромных пространствах образуют хвойные леса, называемые тайгой.
Современные голосеменные представлены преимущественно деревьями, значительно реже — кустарниками и очень редко —лианами; травянистых растений среди них нет. Листья голосеменных значительно отличаются от других групп растений не только по форме и размерам. У большинства видов они игловидные (хвоя) или чешуевидные; Листья располагаются поодиночке, по два или несколько в пучках.
Водопроводящая система (ксилема) голосеменных состоит преимущественно из трахеид, и лишь у некоторых групп имеются настоящие сосуды. Ситовидные трубки не имеют клеток – спутниц .
Древесина имеет камбий, который обеспечивает утолщение.
В древесине находится много смоляных ходов, заполненных смолой.
Подавляющее большинство голосеменных — вечнозеленые, одно- или двудомные растения с хорошо развитыми стеблем и корневой системой, образованной главным и боковым корнями. Расселяются они семенами, которые формируются из семязачатков. Семязачатки голые (отсюда название отдела), Лежат голо на чешуях шишек.
В цикле развития голосеменных наблюдается последовательная смена двух поколений — спорофита и гаметофита с господством спорофита. Гаметофиты сильно редуцированы, причем мужские гаметофиты голо- и покрытосеменных растений не имеют антеридиев, чем резко отличаются от всех разноспоровых бессеменных растений. Представлены пыльцевым зерном с вегетативной и генеративной клетками.
Голосеменные включают шесть классов, два из которых полностью исчезли, а остальные представлены ныне живущими растениями. Наиболее сохранившейся и самой многочисленной группой голосеменных является класс Хвойные,
Семенные растения, как голосеменные, так и цветковые, отличаются от всех остальных представителей растительного мира тем, что размножаются посредством семян, а не с помощью спор.
Что же такое семя и каково его биологическое значение?
все семенные растения, как голосеменные, так и цветковые, являются в то же самое время растениями - разноспоровыми . Это значит, что споры у них двух разных типов - микроспоры (мужские споры) и мегаспоры (женские споры). Первые дают начало мужскому гаметофиту, а вторые - женскому. В то время как процесс оплодотворения у всех бессеменных высших растений происходит в воде, у семенных растений размножение оказалось независимым от воды. Благодаря этому семенные растения могут размножаться половым путем даже в пустыне.
Внутреннее оплодотворение, развитие зародыша внутри семязачатка и появление новой, чрезвычайно эффективной единицы расселения - семени - являются главными биологическими преимуществами семенных растений, давшими им возможность полнее приспособиться к наземным условиям и достигнуть более высокого развития, чем папоротники и другие бессемянные высшие растения .
Преимущества семени над спорой. При размножении спорами каждый раз образуется огромное их число, обычно миллионы, то семян требуется сравнительно небольшое количество. В многоклеточном семени уже содержится зародыш - с корешком, почечкой и зародышевыми листьями - семядолями. Покрыт семенной кожурой - он легко укореняется и начинает самостоятельную жизнь. Кроме того, в семени содержится запас питательных веществ, а спора это одна специализированная клетка.
Голосеменные, как и все семенные растения, являются разноспоровыми. Микроспоры образуются у них в микроспорангиях, а мегаспоры - в мегаспорангиях (нуцеллусах) семязачатков.
Микроспорангии развиваются на микроспорофиллах, а семязачатки - на мегаспорофиллах. Микро- и мегаспорофиллы голосеменных имеют различный внешний вид, размеры и строение. микроспорофиллы и мегаспорофиллы собраны в компактные стробилы (шишки).
У подавляющего большинства голосеменных стробилы однополые, т.е. состоят или только из микроспорофиллов, или только из мегаспорофиллов.
Стробилы, состоящие только из микроспорофиллов, называют мужскими стробилами или мужскими шишками. Стробилы же, состоящие из мегаспорофиллов, носят название женских стробилов или женскими шишиками .
Жизненный цикл голосеменных
Особенности жизненного цикла голосеменных мы рассмотрим на примере сосны обыкновенной
Сосна - крупное дерево, размножающееся семенами. Как известно, семена сосны и других хвойных образуются в шишках. Эти шишки и есть женские стробилы.
Есть мужские шишки (стробилы), в которых созревает пыльца. Они появляются ранним летом и существуют весьма непродолжительное время.
Строение мужского стробила. Микроспорогенез микрогаметогенез
На продольном срезе через мужскую шишку хорошо выделяется ось, к которой крепятся микроспорофиллы . В основании каждого из них размещаются гнезда двух крупных микроспорангиев (пыльцевых мешков) . Гнезда микроспорангиев заполнены археспориальной тканью.
Процесс микроспорогенеза состоит в том, что клетки микроспорангия делятся путем мейоза, в результате образуются тетрады гаплоидных микроспор.
Каждая микроспора (пыльцевое зерно) имеет две оболочки: внутреннюю тонкую - интину и наружную, грубую и прочную - экзину . Характерной особенностью микроспор хвойных являются так называемые воздушные мешки, повышающие аэродинамические качества пыльцы, которая распространяется ветром.
Гаплоидное ядро микроспоры начинает делиться обычным митотическим путем. В результате двух, следующих друг за другом митотических делений, образуется вегетативная и генеративная клетки. – мужской гаметофит.
Из вегетативной клетки развивается пыльцевая трубка. Все дальнейшие изменения происходят с генеративной клеткой. Она делится еще один раз митозом и, в результате этого деления образуются спермии . На этом процесс микрогаметогенеза заканчивается.
Строение женского стробила. Мегаспорогенез и мегагаметогенез
женская шишка (стробил) также имеет ось, к которой крепятся чешуи двух типов: пленчатые кроющие чешуи, в пазухах которых развиваются крупные плодущие или семенные чешуи. В основании семенных чешуй размещается по два семязачатка.
Оба процесса: мегаспорогенез и мегагаметогенез происходят в семязачатке.
Процесс мегаспорогенеза организован очень просто: внутри нуцеллуса клетки макроспорангия делятся мейозом образуется тетрада мегаспор , три из которых быстро погибают и идут на питание единственной оставшейся мегаспоре.
Из гаплоидной мегаспоры митозом образуется женский гаметофит, который со временем занимает практически весь объем семени. По своей функциональной роли женский гаметофит представлен- эндоспермом. Необходимо подчеркнуть, что эндосперм голосеменных растений является гаплоидной тканью, в отличие от триплоидного эндосперма цветковых растений. Гаплоидный эндосперм – запас питательных веществ. На верхнем полюсе гаметофита образуются два типичных архегония. В каждом из них находится крупная яйцеклетка. После формирования яйцеклетки женский гаметофит готов к оплодотворению.
Оплодотворение совершается внутри семязачатка, при этом спермии по пыльцевой трубке проникают к архегониям и один из них сливается с яйцеклеткой. Вторая пара гамет обычно не дает зиготы. Погибает Из зиготы со временем развивается зародыш семени.


Отдел Покрытосеменные (Цветковые ) — отдел высших растений, отличительной особенностью которых является;
1. наличие цветка — увеличивает шансы более эффективного опыления;(опыление насекомыми, ветром,водой)
2.Наличие плода: образование в составе цветка завязи , заключающей в себе семязачатки и предохраняющей их от действия неблагоприятных условий среды; из стенок завязи формируется околоплодник. функция – защита и расселение семян.
3.двойное оплодотворение , в результате которого образуется триплоидный (а не гаплоидные, как у голосеменных) эндосперм; он более жизнеспособный т.к. объединяет признаки отцовского и материнского организмов.
4.редукция гаметофита : мужской гаметофит — пыльцевое зерно состоит из двух клеток: вегетативной и генеративной (генеративная делится, образуя два спермия). Женский гаметофит состоит из 8 клеток зародышевого мешка без архегониев.
5 .Представлены всеми жизненными формами: деревья, кустарники, кустарнички, травы
6 .Ткани высокодифференцированые . Проводящая система хорошо развита. ксилема представлена сосудами, которые более совершенны, чем трахеиды. Ситовидные трубки луба имеют клетки спутницы.
7 . Высокая дифференциация вегетативных органов приспосабливает к различным условиям существования, что позволило покрытосеменным распространится по всему земному шару и занять господствующее положение.
Классы Покрытосеменных (Цветковых)
Выделяют два класса цветковых растений: однодольные и двудольные .
Признаки двудольных растений :
1. Зародыш имеет 2 семядоли,
2. Лист на черешке с сетчатым или ветвящимся жилкованием. Листья простые и сложные
3. Проводящая система состоит из одного кольца проводящих лучков, с камбием; кора и сердцевина дифференцированы.
4. Корневая система стержневая, зародышевый корешок развивается в главный корень.
5. Жизненные формы древесные и травянистые.
6. Цветок имеет число элементов, кратное 5 или 4.
7. околоцветник двойной.(чашелистики и лепестки)
Признаки однодольных растений :
1. Зародыш с одной семядолей, прорастающей подземно,
2. Лист без черешка, с параллельным или дуговым жилкованием, с влагалищным основанием. Листья простые.
3. Проводящая система состоит из многих отдельных пучков; камбий отсутствует; кора и. сердцевина не имеют четкой дифференциации.
4. Корневая система мочковатая, зародышевый корешок рано отмирает, заменяясь системой придаточных корней.
Из вводной главы предыдущего тома мы уже знаем, что проводящая система цветковых растений достигла наиболее высокого уровня эволюционного развития. Проводящая система у цветковых растений оказалась значительно более совершенной, чем у голосеменных (рис. 1), а тем более у папоротников и других групп высших растений. Впрочем, некоторые нз наиболее примитивных представителей ныне живущих двудольных, такие, как виды семейства винтеровых (Winteraceae) и роды троходендрон (Trochodemtron) и тетрацентрон (Tetracentron), по строению проводящей системы мало чем отличаются от примитивных представителей голосеменных типа современных саговниковых или вымерших беннеттитовых. У названных родов нет сосудов, а имеются только трахеиды. В протоксилеме эти трахеиды с кольчатыми и спиральными утолщениями, в метаксилеме обычно лестничные. Бессосудистые формы имеются также среди немногих травянистых двудольных и у ряда однодольных. Конечно, иногда отсутствие сосудов у травянистых растений — явление вторичное (у рясковых).

Как и все другие структурные элементы, членики сосудов в процессе эволюции совершенствуются. Длина их постепенно уменьшается, они становятся шире и в большинстве случаев приобретают более толстые стенки. Сечение их на поперечном срезе становится округлым, лестничные боковые поры заменяются более или менее округлыми окаймленными порами, которые располагаются сначала в горизонтальных рядах (супротивная поровость), а затем расположение их становится очередным, в виде косых рядов (очередная поровость, рис. 2). Возникают ясно выраженные конечные стенки, на первых этапах эволюции еще очень косые. Постепенно они принимают поперечное положение, т. е. располагаются под прямым углом к длине сосуда. По мере укорочения длины и увеличения диаметра члеников сосуда отверстия в лестничной перфорационной пластинке расширяются, число перекладин уменьшается, и в конце концов после исчезновения всех перекладин образуется одна большая перфорация, пазываемая простой (рис. 4). Простая перфорация — это наиболее совершенный тип сквозного отверстия между члениками сосудов, так как сопротивление току жидкости сведено здесь к минимуму. Высшим, наиболее совершенным типом членика сосуда является короткий бочонкообразный членик, ширина которого превосходит его длину (рис. 4). Эволюция члеников сосудов — один из самых ярких и наиболее документированных примеров приспособительной эволюции. Это также пример эволюционного ряда, который завершается кульминационным типом, представляющим собой конечное звено в цепи структурных преобразований.

На более ранних стадиях эволюции ситовидных трубок все ситовидные участки данного членика одинаковы, но затем начинают выделяться участки с более развитыми каллозовыми трубками. Такие, более специализированные ситовидные участки обычно локализуются на определенных стенках члеников, чаще всего на конечных. Части стенки, несущие такие, более специализированные ситовидные участки, называются ситовидными пластинками. Ситовидная пластинка может состоять из нескольких или многих ситовидных участков (с лестничным, сетчатым или иным их расположением). Такая пластинка носит название сложной. Если ситовидная пластинка состоит из одного ситовидного участка, ее называют простой.
Наиболее примитивные формы ситовидных трубок состоят из довольно длинных узких и заостренных, члеников с очень косыми конечными стенками и с более или менее одинаковыми ситовидными участками на конечных и боковых стенках. Подобно конечным стенкам члеников сосудов, конечные стенки члеников ситовидных трубок в процессе эволюции постепенно принимают все менее наклонное положение и в конце концов часто становятся поперечными, т. е. располагаются под прямым углом к боковым стенкам. Одновременно с этим происходит постепенная локализация ситовидных участков на конечных стенках. При этом сложные ситовидные пластинки переходят в простые, более приспособленные для транспорта ассимилятов в растении. Этот процесс аналогичен превращению лестничной перфорации члеников сосудов в простую. В обоих случаях совершенствуется механизм передвижения жидкостей.
Наконец, в процессе эволюции цветковых растений происходило уменьшение длины и увеличение диаметра члеников ситовидных трубок, что, однако, не привело здесь к тем очень коротким и широким бочонкообразным отдельностям, которые встречаются у высокоспециализированных сосудов.
Наблюдается определенная корреляция в эволюции ситовидных трубок и сосудов, и, как правило, уровень специализиции ситовидных трубок соответствует уровню развития сосудов.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Что такое проводящая ткань растений
Проводящие ткани выполняют транспортную функцию, то есть облегчают процесс поглощения питательных веществ растениями. У высших представителей растительности они имеют вид сосудов и ситовидных трубок с пористыми стенками или отверстиями, которые ускоряют проникновение полезных веществ через клетки. Таким образом, в растениях формируется разветвлённая структура, которая объединяет все органы растений воедино. Она тянется от корневой системы до верхушек даже самых листочков и почек.
Проводящими тканями называются ксилема и флоэма, из которых образуется беспрерывная проводящая система. Ксилема – ткани сосудистых растений, которые проводят воду, насыщенную минеральными веществами. Флоэмой называются ткани, которые проводят органические вещества. Они образуются в процессе фотосинтеза.
Особенности проводящей ткани растений
Проводящие ткани имеют сложное строение. Они делятся на множество структур и функциональных элементов. Основными из них являются ксилема и флоэма.
Ксилема состоит из трахей и трахеидов, древесных и паренхиматозных волокон.
Флоэма состоит из проводящих, механических и паренхиматозных тканей. Самые важные – ситовидные трубки и клетки, объединённые в единую систему при помощи межклеточного пространства.
У ксилемы и флоэмы есть некоторые общие особенности:
- их элементы имеют продолговатую форму и вытягиваются по направлению потока веществ;
- поперечные стенки перфорированы и имеют пористую структуру, что благоприятно влияет на проведение питательных веществ;
- проводящие структуры не содержат протопласта, который бы создавал барьер для поступления жидкости;
- оба вида тканей являются сложными, в их состав входят различного рода элементы;
- ксилема с флоэмой объединяются с помощью проводящих пучков.
Роль проводящей ткани
Проводящая ткань выполняет важную функцию, которая состоит в транспортировке питательных веществ, необходимых для полноценного роста и развития растений. Она является необходимой структурной составляющей органов вегетативной системы.
Ткань состоит из комплекса клеток и межклеточного пространства.
Признаки проводящей ткани
В структуру проводящих тканей входят живые и мёртвые продолговатые клетки в форме трубочек.
Стебельки растений имеют пучки проводящих тканей.
Они оснащены сосудами и ситовидными трубками.

Типы и виды проводящей ткани
Проводящие ткани делятся в зависимости от происхождения и периоду образования в организме растений. Если они возникли из первичной васкулярной латеральной меристемы (или прокамбия), тогда они считаются первичными, а которые образовались из вторичной меристемы (или камбия) – считаются вторичными.
Проводящие ткани делятся на два типа:
У них есть общие элементы – ситовидные трубки и сосуды, благодаря которым происходит поток минеральных и органических компонентов.
Проводящая ткань ксилема (древесина)
Ксилема является сложной тканью. В зависимости от функций, в её состав входят:
- трахеальные компоненты, выполняют функцию проводника;
- древесные волокна, отвечают за опорную функцию;
- паренхимные компоненты, выполняют запасающие функции и транспортировку по растению пластических веществ.
Трахеальные элементы
Трахеальные элементы являются самыми высокоспециализированными клетками ксилемы. Они имеют вытянутую форму и как только достигают зрелости – погибают. Их лингнифицированные оболочки покрыты порами и имеют вторичное утолщение.
Процесс вторичные оболочки у них образуются в период роста клетки. В этот период клетки приобретают характерную вытянутую форму. Она может образоваться только до уплотнения оболочки. Для этого у высших растений вырабатывается оптимальное приспособление – вторична оболочка, которая не покрывает клетку полностью. Она располагается в виде колец или спиралек. Благодаря таким уплотнениям молодые трахеальные элементы получаются вытянутыми, не теряя своего объёма.
Несмотря на все достоинства, спиралевидные и кольчатые элементы не отличаются высокой механической прочностью. После достижения пика своего развития, в клетках формируются трахеальные участки со сплошными древесными оболочками. Как только формирование сплошной оболочки заканчивается, клетки прекращают свою жизнедеятельность.
Оболочка тканей, которые отвечают за проведение воды, не бывает идеально ровной. Она имеет пористую структуру с множественными отверстиями. Благодаря этому свойству взрослые структурные элементы получили ещё одно название – точечно-поровые. В результате процесса индивидуального роста и развития имеет место взаимное превращение таких трахеальных структур различной формы (спиральных, сетчатых, лестничных, точечно-поровых).
Проводящие элементы делятся на трахеиды и сосудистые членики. Трахеиды не имеют сквозных отверстий, а для члеников характерно перфорированное дно, что облегчает продвижение жидкости по сосудам.

Первыми обладают высшие растения с давних времён. Они имеют замкнутую оболочку, что затрудняет проникновение воды.
Впоследствии образовались более практичные элементы – членики сосудов. Они имеют на концах каждой клетки отверстия. Из множества таких клеток образуются сосуды, которые свободно переносят воду.
Именно наличие лигнина в их составе позволяют элементам выполнять барьерную функцию.
Трахеальные элементы в процессе развития направлены на защите и транспортировке организма.
Хвощевые, голосеменные и папоротниковые растения имеют древесину с гомогенной ксилемой. В её состав входят преимущественно трахеиды и в небольшом количестве древесные паренхимы. Широколиственные трахеиды с тонкими стенками несут проводящую функцию, а толстостенные выполняют защитную (механическую) функцию.
Покрытосеменные растительные организмы имеют совершенную гетерогенную древесину. В её состав входят сосуды, трахеиды, волокна – либриформы и паренхима, которая имеет свойство запасать питательные вещества.
Сосудистые членики растений имеют разнообразную морфологическую структуру.
Их первначальный эволюционный ряд начинается с трахеид со ступенчатой пористостью и скошенными кончиками. Простейшие лестничные имеют много перегородок. Со временем клетки становятся короче и шире, а их стенки вытягиваются и становятся из скошенного состояния в поперечное состояние.
В самых простых члениках перфорационная стенка состоит из множества перегородок. Со временем они теряются, и образуется одно большое отверстие.
Особенностью цветковых растений является то, что с появлением сосудов, у них сохранились трахеиды, так как совершенство высших растений не является безоговорочным преимуществом. Поэтому во влажной среде скорость проведения воды не особо важна. В них высокий процент члеников сосудов, у сосудов которых лестничная перфорация значительно выше, чем у растительности, растущей в засушливом климате. Соотношение проводящих элементов у различных растений зависит от условий, в которых они находятся, и напрямую определяет их водный баланс.
Второму эволюционном ряду развития проводящих тканей присуща механическая прочность. Трахеиды заменяют волокна либриформа. Это сопровождается уплотнением клеточной оболочки, сужение полости и повышение редукции окаймления пор. А поры между волокнами сузились, приобрели щелевидную форму. Также значительно уменьшилось их количество.
Для паренхимных клеток, из которых состоит древесина, характерным является свойство запасания жиров крахмальных отложений и прочих эргастических веществ.
По мере развития растений у них из латеральной меристемы прокамбия образуется первичная проводящая ткань – ксилема. Некоторым высшим растениям присуща активная работа вторичной боковой меристемы, которая задаёт старт развитию вторичной ксилеме.
В основном, первичную ксилему можно чётко разделить на протоксилему и метаксилему.
Первая стадия – протоксилема – образуется первой. Она состоит только из трахеальных элементов, погружённых в паренхиму. Оболочки таких структур имеют кольчатые утолщения и возможность растягиваться. У метаксилемы ткань имеет сложную организацию. Кроме трахеальных компонентов у неё располагаются волокна, а оболочки намного мощнее. Граница между стадиями роста не может быть чёткой.
Проводящая ткань флоэма (луб)
Флоэмой называется ткань сосудистых растений, которая отвечает за транспортировку пластических веществ, образовавшихся в процессе фотосинтеза от верхушки кроны к корням и по веточкам к плодам или цветкам растений. Как и у ксилемы, клетки флоэмы делятся на несколько типов. Ткань может быть первичной и вторичной. Для первичной флоэмы источником является прокамбий, а для вторичной флоэмы – камбий.
У первичной и вторичной флоэмы есть одинаковые типы клеток:
- ситовидные элементы – состоят из ситовидных клеток или члеников ситовидных трубочек вместе с сопутствующими клетками, которые обеспечивают движение питательных веществ;
- волокна и склереиды – выполняют опорную функцию;
- паренхимные клетки – выполняют запасающую и транспортирующую функцию.
Элементы флоэмы за всё время своего развития перенесли ряд изменений, касающихся их строения и функций.
Проводящие пучки
Проводящими тканями образуются так называемые пучки. К ним примыкает дополнительная ткань, которую называют склеренхимой. Образовавшиеся пучки называются сосудисто-волокнистыми либо армированными.
В зависимости от способности к утолщению пучки бывают открытыми или закрытыми.
Открытые способны к дальнейшему утолщению и образуют камбий. А в закрытых пучках невозможно образование камбия. Также они не могут утолщаться.
У неполных проводящих пучков содержится только один вид ткани (или только флоэма, или только ксилема).
Полные пучки имеют разнообразную конструкцию:
- коллатеральные пучки – флоэма находится над ксилемой;
- биоколлатеральныепучки – имеют дополнительный слой флоэмы;
- концентрические пучки– для них характерно взаимное окружение.
Амфивазильныая ксилема находится вокруг флоэмы, а амфикрибральная ксилема, наоборот, находится внутри флоэмы.
Проводящая ткань жилка
Жилки листа состоят из проводящей ткани. Сосуды ксилемы занимают верхнюю часть, а трубчатая флоэма располагается внизу. Мякоть листа не соприкасается с сосудистыми пучками, которые покрыты плотным слоем клеток паренхимы. Они не имеют в своём составе хлорофилл. Опытным путём доказано, что продукты фотосинтеза из губчатого ткани мезофилла попадают в клетки обкладки, которые перемещают их к ситовидным трубкам.
Также в состав жилки, кроме проводящей ткани, входят механические ткани. Они представлены лубяными и древесинными волокнами, которые обеспечивают прочность и устойчивость листовой пластины.
Функции проводящей ткани
Ксилема выполняет транспортную функцию. Она проводит воду и минеральные вещества, начиная с корневой части и заканчивая плодами и цветками растения. Моховидные растения проводящих элементов не имеют. Их работу компенсируют другие клетки стебля.
В качестве проводящих элементов у многих растений (папоротниковидных, голосеменных и хвощевидных) присутствуют трахеиды. Эти клетки имеют удлинённую форму с косыми заостренными концами и сквозными. Они расположены друг над другом. Транспортировка влаги и минералов при этом не может происходить с высокой скоростью.
У покрытосеменных растений проводящие элементы ксилемы представлены трахеями. Сосуды представлены цилиндрическими клетками, в которых отсутствуют поперечные перегородок. Они находятся друг над другом и образуют канал, через который быстро и без препятствий проходят вода и все минеральные компоненты.
Сосудистые стенки и трахеиды одревеневшие, благодаря этому состоянию обеспечивают прочность всех органов растений. Ксилема имеет не только проводящие свойства, но и создаёт механическую ткань – склеренхим, которая выполняет опорную и запасающую функции.
Флоэма отвечает за транспортировку органических элементов от листочков вниз к корневой системе. Это осуществляется с помощью ситовидных трубочек. Строение клеток имеет свои особенности: наличие цитоплазмы и отсутствие ядра. Их цитоплазмы сообщаются благодаря мелким отверстиям в поперечных стенках, которые визуально напоминают сито.
Покрытосеменные растения вместе с ситовидными трубками клетки имеются сопутствующие клетки с ядрами, которые способны исполнять дополнительные функции.
К флоэме относится также механическая (склеренхима) и запасающая ткань (паренхима). Вместе с проводящей системой ксилемы и флоэмы с волокнами паренхимы образуются пучки из сосудов и волокон, которые проникают во все органы растений.
Строение проводящей ткани
Проводящая ткань имеет живые или мёртвые удлинённые трубковидные клетки. Она сосредоточена в стеблях и листочках растений. В её составе выделяют сосуды и ситовидные трубки. Сосудами называются длинные трубки из отмерших клеток, которые не имеют перегородок. Они являются проводниками воды и минеральных питательных веществ. Именно по ним из корней по стеблям питание поступает к верхушкам.
Ситовидные трубки являются пористыми живыми клетками, без наличия ядра. Через них питательные вещества попадают из листьев к другим частям растений.

На спилах деревьев хорошо выделяются луб и древесина. Древесный слой представлен сосудиками, которые проводят вверх питательные вещества, а луб имеет трубочки, по которым питательные вещества спускаются от верхней части к корням.
Клетки проводящей ткани
У проводящей ткани имеются трахеиды и членики сосудов Трахеиды имеют вытянутую форму с целостными стенками. Транспорт необходимых для роста и развития компонентов происходит благодаря пористой структуре.
Членики сосудов являются вторым важным проводящим элементом. Они находятся друг над другом, и в местах их соприкосновения образуются пространство, которое называется перфорацией. Такие промежутки нужны для транспортировки полезных веществ по всем сосудам растения. В отличие от трахеид, скорость перемещения по сосудам значительно выше.
У обоих проводящих элементов клетки не имеют протопластов, тог есть признаком живых клеток. Благодаря этому транспортировка полезных веществ происходит быстро и без преград. Сосуды и трахеиды переносят растворы не только по вертикали, но и по горизонтали.
Прочная структура клеток обусловлена наличием уплотнению на стенках клеток. По типу утолщения на проводящих элементах бывают спиральными кольчатыми, лестничатыми, сетчатыми и сетчато-поровыми.
Проводящая ткань корня
Проводящие ткани располагаются в корневой системе и побегах. Они состоят из ксилемы и флоэмы. Благодаря им у растений проходит восходящий и нисходящий ток полезных веществ
Восходящий ток происходит благодаря ксилеме, по которой вверх поднимаются вода и минеральные компоненты.
Нисходящий ток происходит благодаря флоэме. Она обеспечивает транспорт органических веществ, которые синтезируются в надземных частях растения и спускаются вниз.
Проводящая ткань листа
Проводящие ткани образуют основу листьев, которая получила название жилка листа. В их состав входит первичные формы ксилемы и флоэмы, производных из прокамбия, которые объединяются в закрытые коллатеральные сплетения (пучки). Благодаря им в листочке образуется непрерывная система, которая напрямую связана с проводящей системой стебля растения. Только жилки среднего и крупного размера некоторых двудольных растений могут второй раз увеличиваться в ширину. Ксилема в листе направлена на верхнюю плоскость листа, а флоэма – на нижнюю его часть.
Пучки мелких жилок имеют немного проводящих элементов. На кончиках жилок находятся трахеальные элементы. Пучкам не свойственно соприкасаться с мезофиллом листьев. Их защищают крупные обкладочные клетки, которые регулируют движение веществ. Для листьев многих растений характерна связь эпидермы и проводящих пучков некоторыми структурными элементами, которые не только проводят воду, но и укрепляют лист.
Самую важную роль имеют небольшие жилки, находящиеся внутри мезофилла. Благодаря им лист насыщается влагой и всеми необходимыми минералами.
Ксилема выполняет в растении две основные функции: по ней движется вода вместе с растворенными минеральными веществами и она служит опорой органам растения. Таким образом, ксилема играет в растении двоякую роль — физиологическую и структурную. В состав ксилемы входят гистологические элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. На рис. 6.9 эти гистологические элементы представлены и поперечном и продольном разрезах.
Трахеиды ксилемы
На рисунке представлено строение трахеид. У покрытосеменных число трахеид по сравнению с числом сосудов относительно невелико. Сосуды считаются более эффективным приспособлением для транспорта воды, нежели трахеиды; появление сосудов связано, как полагают, с тем, что у покрытосеменных с их большой листовой поверхностью транспира-ция идет более активно.
Сосуды ксилемы
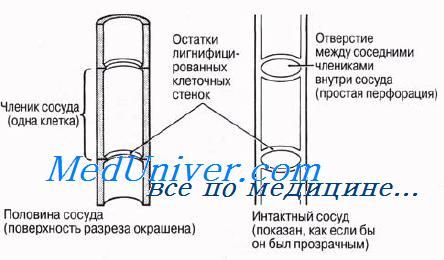
Протоксилема и метаксилема
Первые по времени образования сосуды — протоксилема — закладываются на верхушке осевых органов, непосредственно под верхушечной меристемой, там, где окружающие их клетки еше продолжают вытягиваться. Зрелые сосуды про-токсилемы способны растягиваться одновременно с вытягиванием окружающих клеток, поскольку их целлюлозные стенки еще не сплошь одревеснели —лигнин откладывается в них лишь кольцами или по спирали (рис. 6.12). Эти отложения лигнина позволяют трубкам сохранять достаточную прочность во время роста стебля или корня. С ростом органа появляются новые сосуды ксилемы, которые претерпевают более интенсивную лигнификацию и завершают свое развитие в зрелых частях органа; так формируется ме-гаксшема. Тем временем самые первые сосуды протоксилемы растягиваются, а затем разрушаются. Зрелые сосуды метаксилемы не способны растягиваться и расти. Это мертвые, жесткие? полностью одревесневшие трубки. Если бы их развитие завершалось до того, как закончилось вытягивание окружающих живых клеток, то они бы очень сильно мешали этому процессу.
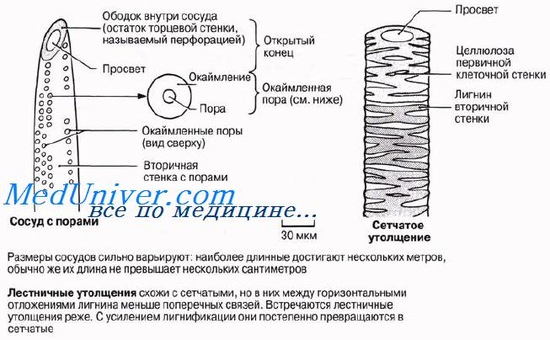
У сосудов метаксилемы обнаруживаются три главных типа утолщений: лестничные, сетчатые и точечные.
Длинные полые трубки ксилемы — идеальная система для проведения воды на большие расстояния с минимальными помехами. Так же как и в трахеидах, вода может переходить из сосуда в сосуд через поры или через неодревеснев-шие части клеточной стенки. Вследствие одревеснения клеточные стенки сосудов обладают высокой прочностью на разрыв, что тоже очень важно, потому что благодаря этому трубки не спадаются, когда вода движется в них под натяжением (разд. 13.4).
Вторую свою функцию — механическую — ксилема выполняет также благодаря тому, что она состоит из ряда одревесневших трубок. В первичном теле растения ксилема в корнях занимает центральное положение, помогая корню противостоять тянущему усилию надземных частей, изгибающихся под порывами ветра, В стебле проводящие пучки либо образуют по периферии кольцо, как у двудольных, либо располагаются беспорядочно, как у однодольных; в обоих случаях стебель пронизывается отдельными тяжами ксилемы, обеспечивающими ему определенную опору. Особенно важное значение опорная функция ксилемы приобретает там, где имеет место вторичный рост. Во время этого процесса быстро нарастает количество вторичной ксилемы; к ней переходит от колленхимы и склеренхимы роль главной механической ткани, и именно она служит опорой у крупных древесных и кустарниковых пород. Рост стволов в толщину определяется в известной мере нагрузками, которым подвергается растение, так что иногда наблюдается дополнительный рост, смысл которого состоит в усилении структуры и обеспечении ей максимальной опоры.
Древесинная паренхима ксилемы
Древесинная паренхима ксилемы содержится как в первичной, так и во вторичной ксилеме, однако в последней ее количество больше и роль важнее. Клетки древесинной паренхимы, подобно любым другим паренхимным клеткам, имеют тонкие целлюлозные стенки и живое содержимое.
Во вторичной ксилеме имеются две системы паренхимы. Обе они возникают из меристемати-ческих клеток, называемых в одном случае лучевыми инициалями, а вдругом — веретеновидны-ми инициалями (гл. 22). Лучевая паренхима более обильна. Она образует радиальные слои ткани, так называемые сердцевинные лучи, которые, пронизывая сердцевину, служат живой связью между сердцевиной и корой. Здесь запасаются различные питательные вещества, скапливаются таннины, кристаллы и т. п., и здесь же осуществляется радиальный транспорт питательных веществ и воды, а также газообмен по межклетникам.
Из веретеновидных инициалей обычно развиваются сосуды ксилемы и ситовидные трубки флоэмы вместе с их клетками-спутницами, однако время от времени они дают начало также и паренхимным клеткам. Эти паренхимные клетки образуют во вторичной ксилеме вертикальные ряды.
Древесинные волокна ксилемы
Полагают, что древесинные волокна, так же как и сосуды ксилемы, ведут свое происхождение от трахеид. Они короче и уже трахеид, а стенки их гораздо толще, но поры их сходны с порами, имеющимися в трахеидах, и на срезах волокна иногда трудно отличить от трахеид, поскольку между теми и другими есть ряд переходных форм. Древесинные волокна очень напоминают уже описанные волокна склеренхимы; их торцевые стенки также перекрываются. В отличие от сосудов ксилемы древесинные волокна не проводят воду; поэтому у них могут быть гораздо более толстые стенки и более узкие просветы, а значит, они отличаются и большей прочностью, т. е. придают ксилеме дополнительную механическую прочность.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: