
Развитие семязачатков у покрытосеменных
Обновлено: 05.10.2024
Семязачаток – это небольшое образование, которое располагается в завязи пестика. Число семязачатков в завязи пестика может быть от одного (пшеница, ячмень, слива, подсолнечник) до нескольких тысяч (мак) и миллионов (орхидные). В семязачатке происходят процессы формирования мегаспор, женского гаметофита и процесс оплодотворения. После оплодотворения он развивается в семя.
Сформированный семязачаток представляет собой многоклеточное образование. К плаценте он прикрепляется семяножкой – фуникулюсом.
Снаружи семязачаток покрыт одним-двумя покровами – интегуметами. На верхушке интегументы не смыкаются, образуя таким образом канал – микропиле (пыльцевход).
Под интегументами расположен многоклеточный нуцеллус, или ядро семязачатка (гомологичен мегаспорангию).
Конец семязачатка, противоположный микропиле, называют халазой. На уровне халазы происходит слияние нуцеллуса и интегументов (рис. 13).
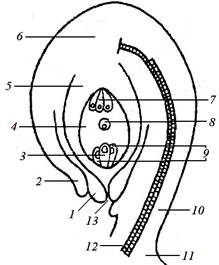
Рис. 13. Строение сформированного семязачатка:
1, 2 – внутренний и наружный интегументы; 3 – яйцеклетка; 4 – зародышевый мешок; 5 – нуцеллус; 6 – халаза; 7 – антиподы; 8 – вторичное ядро; 9 – синергиды; 10 – семяножка;
11 – плацента; 12 – проводящий пучок; 13 – пыльцевход (микропиле)
Особенности строения зрелого семени во многом определяются типом образовавшего его семязачатка. В зависимости от ориентации, взаимного расположения частей и степени изогнутости нуцеллуса различают пять основных типов семязачатков (рис. 14):
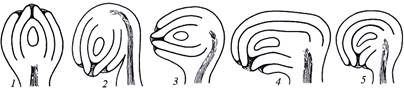
Рис. 14. Основные типы семязачатков:
1 – ортотропный; 2 – анатропный; 3 – гемитропный; 4 – кампилотропный;
1) ортотропный (прямой) – микропиле и семяножка находятся на противоположных концах оси семязачатка (гречишные, ореховые);
2) анатропный (обратный, обращенный) – нуцеллус повернут по отношению к прямой оси на 180°, поэтому микропиле и семяножка расположены рядом (самый распространенный тип семязачатка);
3) гемитропный (полуповернутый) – семязачаток повернут на 90°, в результате микропиле и нуцеллус расположены по отношению к семяножке под углом 90° (некоторые первоцветные, норичниковые);
4) кампилотропный (односторонне изогнутый) – нуцеллус изогнут только микропилярным концом, фуникулюс и микропиле расположены рядом (бобовые, мальвовые);
5) амфитропный (двусторонне изогнутый) – нуцеллус изогнут с двух сторон, в виде подковы, микропиле и фуникулюс расположены рядом (тутовые, ладанниковые).
Мегаспорогенез – это процесс формирования мегаспор в нуцеллусе семязачатка.В нуцеллусе вблизи микропиле обособляется, увеличивается одна (реже несколько) клета археспория. Эта клетка становится материнской клеткой мегаспор, она имеет диплоидный набор хромосом (2n). Материнская клетка однократно делится мейозом, в результате возникает тетрада гаплоидных мегаспор (четыре по 1n). Мегаспоры располагаются линейно от микропиле к халазе (рис. 15).
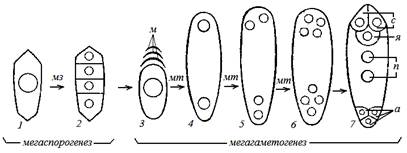
Рис. 15. Развитие зародышевого мешка (схема):
1 – материнская клетка; 2 – тетрада мегаспор; 3 – одна развитая мегаспора и три отмирающие; 4 – двухъядерный мешок; 5 – четырехъядерный мешок; 6 – восьмиядерный мешок; 7 – развитый зародышевый мешок; с – синергиды; я – яйцеклетка;
п – полярные ядра; а – антиподы; м – три неразвитые мегаспоры; мз – мейоз;
Мегагаметогенез – это процесс формирования женского гаметофита, которым является восьмиядерный зародышевый мешок. Одна из четырех мегаспор, которая находится ближе к халазе, делится быстрее. Поэтому остальные три мегаспоры подавляются в развитии, т. е. дегенерируют.
Единственная мегаспора увеличивается, разрастается, ее ядро трехкратно делится митозом. В результате в сильно растянутой клетке мегаспоры формируется восемь ядер, которые располагаются по четыре на халазальном и микропилярном полюсах. Далее происходит дифференцировка ядер: от полюсов к центру отходит по одному ядру. Эти ядра называют полярными. Оставшиеся ядра обособляются в клетки. На микропилярном полюсе центральная крупная клетка преобразуется в яйцеклетку, две рядом расположенные боковые клетки – в синергиды. Яйцеклетка с синергидами образует яйцевой аппарат. На противоположном халазальном полюсе три клетки образуют группу антипод. В центре два полярных ядра сливаются, в результате образуется вторичное (центральное) ядро зародышевого мешка (см. рис. 13).
Таким образом, единственная мегаспора, прорастая внутри семязачатка, образует женский гаметофит – зародышевый мешок. Он включает:
а) яйцевой аппарат состоит из яйцеклетки и двух синергид; они расположены на микропилярном полюсе семязачатка;
б) три антиподы – расположены на халазальном полюсе семязачатка; яйцеклетка, синергиды и антиподы являются гаплоидными (1n);
в) вторичное (центральное) ядро зародышевого мешка – образовано при слиянии двух полярных ядер; оно диплоидное (2n).

Семязачаток Escallonia rubra: 1 – микропиле; 2 – фуникулус; 3 – интегумент; 4 – интегументальный тапетум; 5 – халаза; 6 – нуцеллус; 7 – зародышевый мешок.
СЕМЯЗАЧА́ТОК (семяпочка), орган, развивающийся в завязи покрытосеменных и в стробилах (шишках) голосеменных растений; зачаток семени. Состоит из нуцеллуса (мегаспорангия), окружённого одним или двумя интегументами (покровами), которые, смыкаясь, формируют микропиле (пыльцевход), а также противоположной ему части С. – халазы и фуникулуса (семяножки), с помощью которого С. прикрепляется к плаценте. В нуцеллусе С. дифференцируется археспорий, осуществляются мейоз, развитие мегаспор, женского гаметофита и происходит оплодотворение.
Прежде чем перейти к строению семязачатка, рассмотрим его основные морфологические типы (рис. 25), установленные еще французским ботаником Ш. Мирбезем в 1829 г.


Рис. 25. Различные формы семязачатков:
1 - ортотропный; 2 - анатропный: 3 - гемитропный; 4 - кампилотропный; 5 - амфитропный.
У многих цветковых растений микропиле семязачатка находится на одной оси с семяножкой (фуникулусом) и, таким образом, семязачаток расположен перпендикулярно к плаценте. Такой радиально-симметричный семязачаток был назван ортотропным (от греч. orthos — прямой, правильный и tropos — поворот, направление) или прямым. (рис. 25). Нередко его называют атропным (от греч. а — частица отрицания). Ортотропные семязачатки характерны для хлорантовых, савруровых, перцевых, роголистника, платана, мириковых, ореховых, валлиснерии, взморника, рдеста, ксирисовых, эриокауловых, рестиевых и некоторых других родов и семейств.
Гораздо более распространены семязачатки, загнутые вниз (в сторону плаценты) на 180° и приросшие вследствие этого своим боком к удлиненной семяножке. Поэтому микропиле у них обращено к плаценте и расположено рядом с основанием семязачатка. Такие семязачатки называются анатропными (от греч. anatrope — перевернутый, обращенный) или обращенными (рис. 25, 26). Анатропные семязачатки характерны для большинства цветковых, как двудольных, так и однодольных. Известный чешский ботаник Ф. Нетолицкий (1926) пришел к выводу, что анатропный тип является исходным, а ортотропный — производным, что вполне согласуется с современными представлениями.
В тех случаях, когда семязачаток вследствие более сильного разрастания одной стороны становится изогнутым, он называется кампилотропным (от греч. kampylos — изогнутый) или полусогнутым . В отличие от анатропного и гемитропного типов здесь изогнуты интегумент и даже мегаспорангий (нуцеллус). Этот тип семязачатка характерен для многих представителей порядка гвоздичных, для каперсовых, некоторых бобовых, для филезиевых, некоторых злаков и ряда других групп. В большинстве случаев кампилотропные семязачатки возникают из анатропных, но не исключено, что иногда они могут возникать и из ортотропных.
В некоторых случаях семязачаток перегибается посередине таким образом, что в продольном разрезе его мегаспорангий имеет подковообразную форму. Это амфитропный (от греч. amphi — оба) семязачаток. Он встречается у лейтнерии, кроссосомы, кнеоровых некоторых акантовых и немногих других растений (рис. 25).
Между всеми этими типами имеются промежуточные формы, и поэтому очень часто бывает трудно определить, какой тип семязачатка имеет то или иное растение. Тем не менее выделение этих типов оказалось очень полезным, так как они имеют определенное систематическое значение.
Как это уже давно указывал датский ботаник Й. Э. Варминг (1913), в систематическом отношении важно также, куда загнуты анатропные или кампилотропные семязачатки — к верхушке (в этом случае они называются эпитропными) или ко дну завязи (они носят название апотропных; рис. 27).

Читатель может спросить: почему мегаспорангий семязачатка называют нуцеллусом, если известно, что это мегаспорангий? Дело в том, что долгое время это не было известно, пока в 1851 г. великий немецкий ботаник В. Гофмейстер не установил, что образование, давно известное под названием нуцеллуса, представляет собой не что иное, как мегаспорангий. Но, как часто бывает в науке, даже после замечательных исследований Гофмейстера мегаспорангий семенных растений продолжали называть нуцеллусом, а некоторые ботаники, как, например, американский морфолог А. Имс (1961), подвергали сомнению даже само наличие мегаспорангия у цветковых растений.
Однако большинство ботаников справедливо считают, что наличие у цветковых мегаспорангия, правда очень упрощенного и редуцированного, не вызывает сомнений. Ведь в нуцеллусе имеются два необходимых элемента каждого спорангия — спорогенная ткань (обычно крайне редуцированная) и стенка, которая, однако, гистологически сильно упрощена. Совершенно очевидно, что нуцеллус цветковых морфологически вполне соответствует нуцеллусу голосеменных, т. е. они представляют собой гомологичные образования и являются мегаспорангиями.
Как и у голосеменных, мегаспорангий цветковых окружен интегументом. Однако, в то время как у голосеменных имеется лишь один интегумент, который, согласно гипотезе Маргариты Бенсон (1904), произошел из обеспложенного периферического слоя спорангиев, окружавших центральный фертильный спорангий, у подавляющего большинства цветковых имеются два интегумента. В связи с этим различают битегмальные (от лат. bis — дважды и tegmen — покров) семязачатки, т. е. семязачатки с двумя интегументами, и унитегмальные (от лат. unus — один), т. е. семязачатки с одним интегументом.
В равных линиях эволюции цветковых растений из битегмальных семязачатков произошли унитегмальные. Унитегмальные семязачатки характерны для березовых, мириковых, ореховых, вересковых и близких им семейств, кизиловых, аралиевых, зонтичных, для различных семейств двудольных, характеризующихся спайнолепестным венчиком, и для некоторых однодольных. По-видимому, в большинстве случаев унитегмальные семязачатки произошли из битегмальных в результате слияния обоих покровов, но в ряде случаев они возникли вследствие недоразвития внешнего или чаще внутреннего покрова. Так, в семействе раффлезиевых наблюдается редукция внешнего интегумента, а у рода митрастемон (Mitrastemon) он совершенно подавлен и семязачатки унитегмальные. У родственного семейства гидноровых семязачаток постоянно унитегмальные. В то же время у некоторых представителей семейства розовых, таких, как гравилат, земляника, лапчатка и манжетка, унитегмальный семязачаток возник в результате недоразвития внутреннего покрова. В некоторых семействах, таких, как лютиковые, розовые, бобовые, ивовые, пальмы, орхидные и другие, даже близкие роды отличаются числом интегументов. Это показывает, что унитегмальные семязачатки возникли из битегмальных независимо в самых разных ветвях развития цветковых растений.
У некоторых паразитных двудольных покров семязачатка исчезает полностью и мегаспорангий оказывается, таким образом, голым. Такие атегмальные (от греч. а — частица отрицания) семязачатки характерны для многих, представителей порядка санталовых. У некоторых баланофоровых утрачен не только покров семязачатка, но даже стенка мегаспорангия.

Рис. 27. Типы анатропного семязачатка в зависимости от его поворота в гнезде и от направления фуникулуса: в левом ряду - эпитропные, в правом - апотропные.
В семязачатках наблюдаются, два типа строения мегаспорангия и в соответствии с этим различают два типа семязачатков, названные Ф. Ван Тигемом (1898) крассинуцеллятными (от лат. crassus — толстый и nucellus — ядрышко) и тенуинуцеллятными (от лат. tenuis — тонкий). Крассинуцеллятные семязачатки характеризуются относительно мощно развитым мегаспорангием, и поэтому материнская клетка мегаспор (мегаспороцит) отделена от эпидермы мегаспорангия одним или несколькими слоями клеток. У тенуинуцеллятных семязачатков, мегаспорангий которых более редуцирован, материнская клетка мегаспор лежит непосредственно под эпидермой мегаспорангия. Эти два типа не всегда бывают строго разграничены, и между ними известны различные переходы. Первый тип является более примитивным, второй тип возник из первого в результате редукции мегаспорангия. Поэтому крассинуцеллятные семязачатки характерны для относительно более примитивных двудольных и для большинства однодольных, в то время как тенуинуцеллятные семязачатки встречаются у относительно более подвинутых двудольных и лишь у немногих однодольных. Кроме того, крассинуцеллятные семязачатки чаще всего бывают битегмальными, а тенуинуцеллятные, как правило, унитегмальные.
У цветковых растений в мегаспорангии образуется обычно одна материнская клетка мегаспор. Она претерпевает два деления мейоза, в результате чего аналогично появлению тетрады гаплоидных микроспор образуется тетрада гаплоидных мегаспор. Как и при формировании микроспор, появляются каллозовые оболочки, обеспечивающие изоляцию материнской клетки и мегаспор от окружающих клеток.
Мегаспоры тетрады бывают расположены по-разному, но чаще всего образуется линейная тетрада, реже — Т-образная и еще реже — квадратная или тетраэдрическая. У громадного большинства цветковых растений возникают четыре мегаспоры, из которых нижняя (халазальная) и значительно реже верхняя (микропилярная), или промежуточная, увеличиваются в размерах и функционирует как мегаспора, а остальные три отмирают и разрушаются. Очень редко вместо четырех потенциальных мегаспор образуются три, две, а иногда только одна. В последнем случае материнская клетка мегаспоры становится непосредственно мегаспорой.
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру: от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.

Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) - важнейшими производителями органических веществ на суше, как водоросли - в морях и океанах.
Цветок - генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения. Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище - завязи, сформированной из плодолистика (-ов).
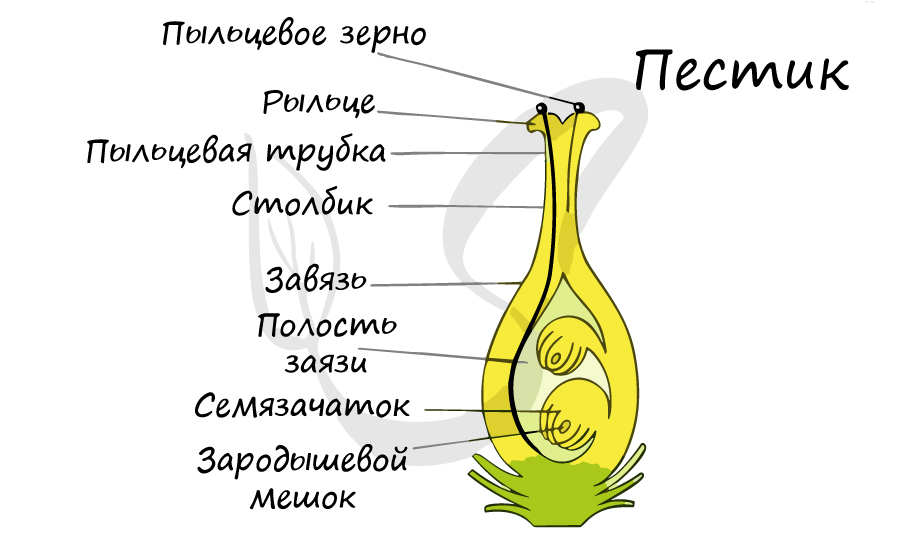
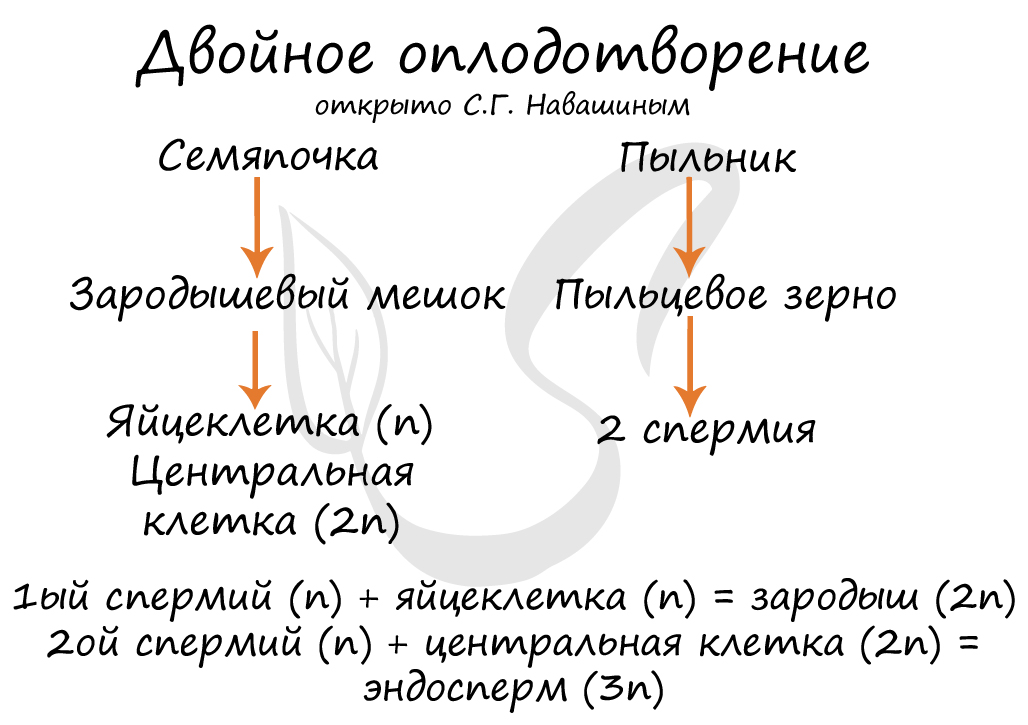
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества - эндосперма (3n). Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод - генеративный орган, служащий для защиты и распространения семян.
Ксилема - проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами, а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

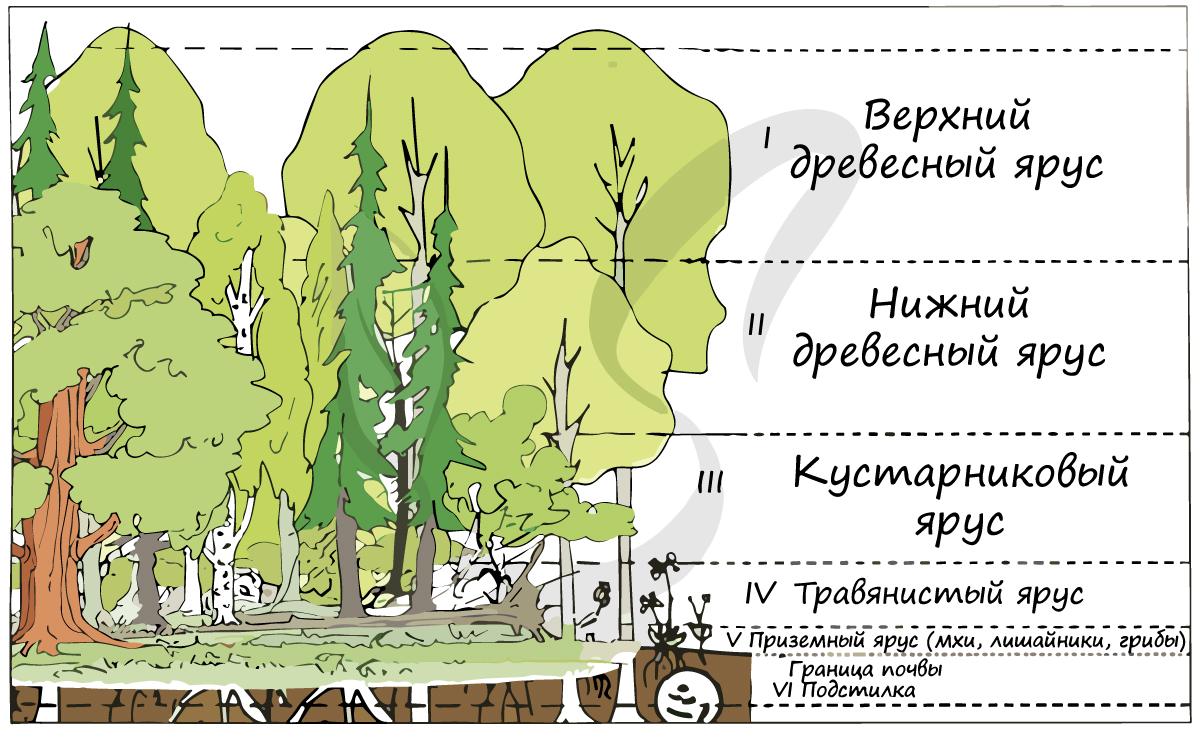
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых :)
Классы покрытосеменных
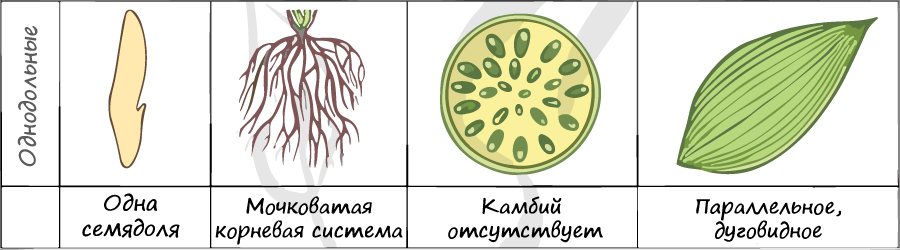
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные, бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
-
Двудольные - семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
-
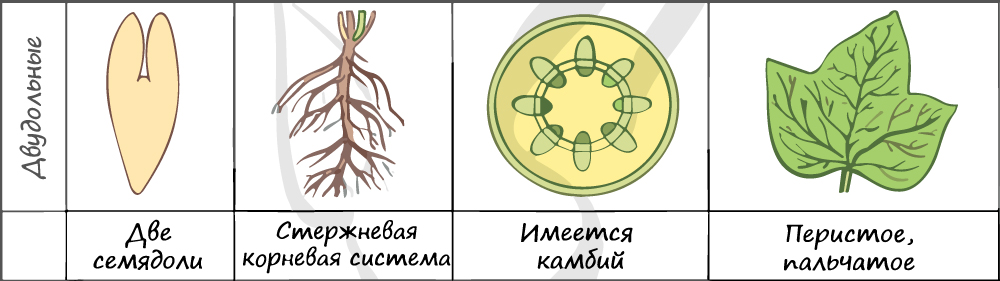
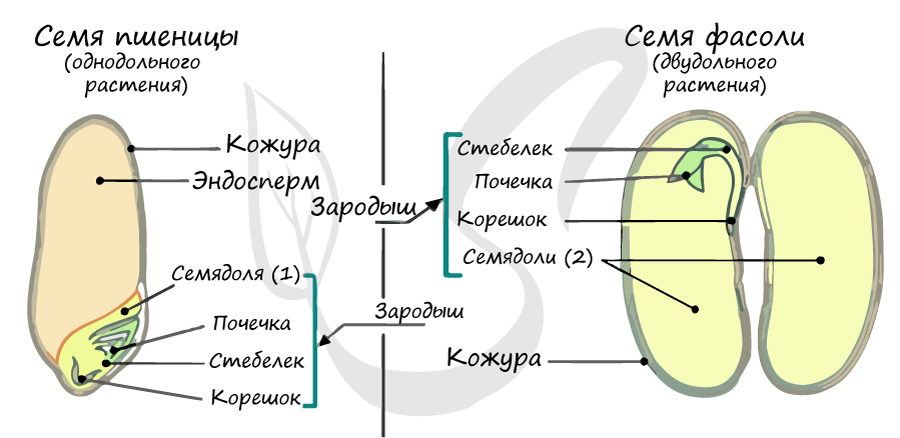
В составе зародыша обычно имеется две семядоли
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни. Главный корень развивается из зародышевого корешка.
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

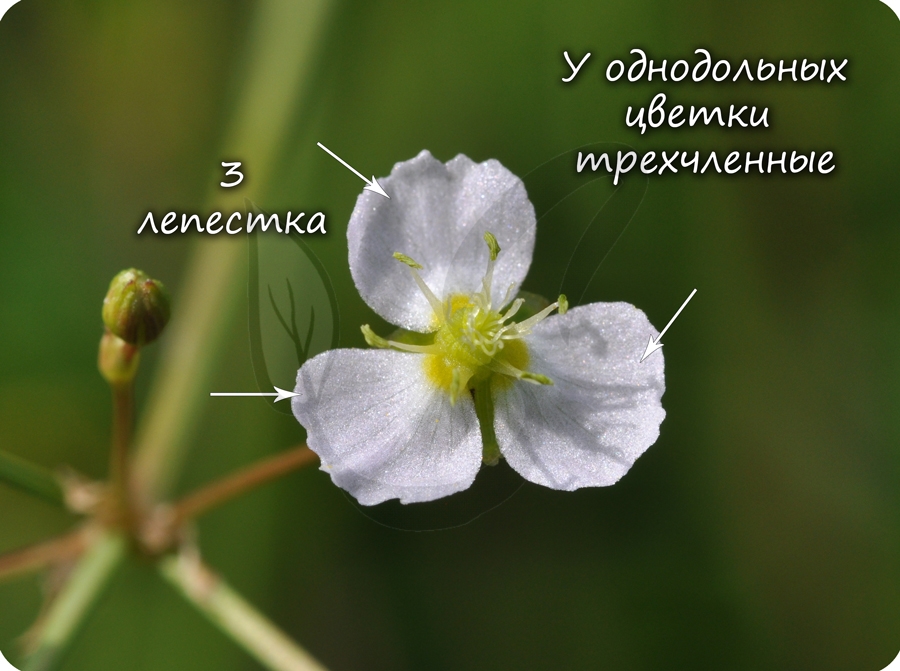
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
Эндосперм семени
Эндосперм (от греч. endon - внутри + греч. sperma - семя) - запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у - березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
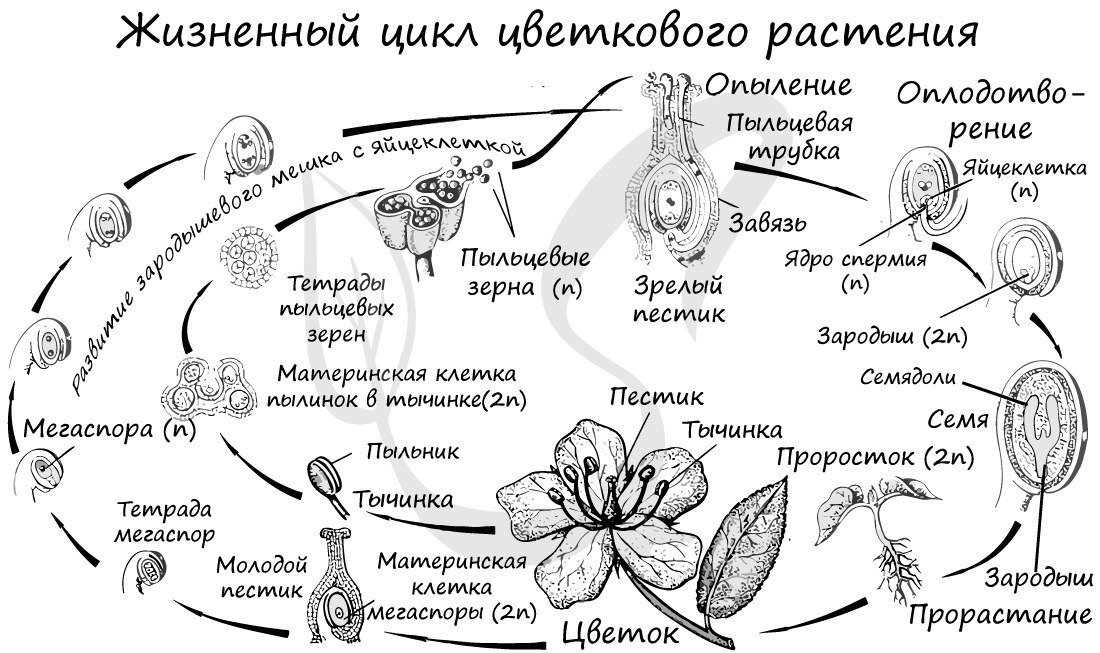
Жизненный цикл
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n) в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) - мужской гаметофит. В завязи пестика в семязачатке формируется женский гаметофит - зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток. Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) - запасного питательного вещества. Другой спермий (n) сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник - образуется плод. Своим внешним видом плоды привлекают животных, и те их охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия, они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте - почти все культурные растения принадлежат к этому отделу! Цветковые имеют медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: