
Самоопыление затруднено или невозможно когда выберите три правильных ответа
Обновлено: 05.09.2024
При перекрестном опылении пыльца с одного растения попадает на другое растение. Таким образом, в образовании нового организма участвуют два родительских растения. У перекрестноопыляемых растений существуют специальные приспособления, препятствующие самоопылению. Например, тычинки и пестики могут быть разной длины. Тычиночные и пестичные цветки на одном растении могут раскрываться в разное время. У двудомных растений самоопыление невозможно в принципе.
При перекрестном опылении нужен какой-нибудь посредник, который бы перенес пыльцу с одного цветка на другой. Часто таким посредником выступают насекомые. Цветки привлекают их своим сладким нектаром, приятным запахом, яркими лепестками. Насекомое садится на цветок, к его лапкам прилипает пыльца. Когда насекомое перелетит на другой цветок, то пыльца может попасть на пестик. Таким образом произойдет опыление.
Различают несколько основных типов опыления:
1 . Самоопыление
2 . Перекрёстное опыление
3 . Искусственное опыление.
Теперь поговорим о них поподробнее:
1 ). При самоопылении пыльца из тычинки попадает на рыльце пестика того же самого цветка или другого цветка того же растения.
Так опыляются пшеница, рис, овёс, ячмень, горох, фасоль, хлопчатник. Самоопыление у растений чаще всего происходит в ещё не раскрывшемся цветке, т. е. в бутоне. Когда цветок раскроется, оно уже закончено. При самоопылении сливаются половые клетки, образовавшиеся на одном растении и, следовательно, имеющие одинаковые наследственные признаки. Вот почему потомство, образовавшееся в результате самоопыления, очень похоже на родительское растение.
2 ). При перекрёстном опылении пыльца с тычинок цветка одного растения переносится на рыльца пестиков цветков других растений.
При перекрёстном опылении происходит перекомбинация наследственных признаков отцовского и материнского организмов, и образовавшееся потомство может приобрести новые свойства, которых не было у родителей. Такое потомство более жизнеспособно. Перекрёстное опыление встречается значительно чаще, чем самоопыление. Более того, у растений выработались специальные приспособления, затрудняющие самоопыление или делающие его невозможным: тычинки и пестики на одном цветке созревают неодновременно, тычинки бывают короче пестиков. У таких растений, как кукуруза, огурец, арбуз, орешник, самоопыление затруднено, потому что тычинки и пестик и у них находятся в разных цветках. Невозможно самоопыление и у двудомных растений (тополь, ива). У них мужские и женские цветки расположены на разных растениях. Вы знаете, что у голосемянных растений пыльцу переносит ветер. У покрытосеменных её могут переносить, помимо ветра, насекомые, птицы, вода.
3 ). Также иногда в роли опылителя выступает человек. При этом он иногда сознательно переносит пыльцу с тычинок на рыльца пестиков. Такое опыление называют искусственным. Его осуществляют с целью выведения новых сортов и повышения урожайности некоторых растений. Переносят пыльцу чистой, сухой, мягкой кисточкой или кусочком резины, прикреплённым к проволоке. У некоторых растений цветки готовят к опылению заранее, пока они не распустились. Для этого осторожно открывают бутоны и удаляют из них тычинки, чтобы не произошло самоопыление.
Затем на бутоны надевают марлевые мешочки, чтобы ветер или насекомые случайно не занесли пыльцу на рыльца. Когда эти бутоны распустятся, на их рыльца наносят заблаговременно заготовленную пыльцу.
Нашли ошибку?
Если Вы нашли ошибку, неточность или просто не согласны с ответом, пожалуйста сообщите нам об этом
Возможность самоопыления у цветковых растений с обоеполыми цветками отнюдь не исключена, несмотря на наличие у них разнообразных устройств, противодействующих ему. Для подавляющего большинства покрытосеменных, за исключением двудомных и строго самонесовместимых видов, следует допустить, что некоторая, иногда довольно большая, часть семян ежегодно образуется в результате самоопыления.
Наряду со случайной автогамией нередко встречается более или менее регулярная автогамия, возникающая под давлением условий, затрудняющих перекрестное опыление или препятствующих ему. Это вынужденное самоопыление. Оно представляет резервный способ опыления и должно рассматриваться как адаптация к неблагоприятным условиям среды.
Самоопыление у однолетних растений встречается чаще, чем у многолетних. Однолетние самоопыляющиеся виды имеются во многих семействах (крестоцветные, бобовые, злаки и др.). Примерами их могут служить крошечные мелкоцветковые растения резушка Таля(Arabidopsis thialiana), пастушья сумка, веснянка весенняя, клевер скученный, клевер шершавый, клевер подземный, герань нежная и др. Они растут на сухих и бедных почвах, быстро завершают жизненный цикл, самоопыление у них очень обычно и связано с особым: образом жизни. Ему благоприятствуют, видимо, два обстоятельства. Прежде всего популяции однолетних растений подвержены сильным колебаниям численности в разные годы. Самоопыление позволяет каждой такой популяции легче и быстрее восстановиться после понесенного ею ущерба, когда она уменьшается до малого числа особей или даже до единственного индивидуума. Кроме того, однолетние растения обычно связаны с узко ограниченными и специфическими местообитаниями. Благодаря самоопылению адаптация этих однолетних видов к таким условиям оказывается вполне устойчивой, что дает им известные преимущества в борьбе за жизнь. Здесь напрашивается аналогия с однолетними самоопыляющимися культурными растениями (культурные виды пшениц, ячменя, овса, гороха посевного и др.), гомозиготные популяции которых в определенных условиях культуры вполне устойчивы и продуктивны.
Самоопыление возникает на базе ксеногамного опыления в результате нарушения (дегенерации) важнейших его механизмов — самонесовместимости и дихогамии. Вследствие подавления генов самонесовместимости происходит сдвиг в сторону более или менее полной самосовместимости. Дихогамия становится как бы стертой вследствие удлинения начальных фаз (рыльцевой и тычиночной) в развитии цветка, благодаря чему эти фазы совмещаются или налегают друг на друга.
Автогамия происходит в обоеполых цветках в разные периоды цветения — в самом начале его, .иногда еще в бутонах (бутонная автогамия), на протяжении всего цветения или в самом конце его.
Бутонная автогамия встречается редко. Более обычной является автогамия в конце периода цветения, когда перекрестное опыление с помощью ветра или насекомых по каким-либо причинам, чаще из-за дождливой и холодной погоды или из-за отсутствия опылителей, не осуществилось. В этом случае страхующая функция автогамии выступает очень наглядно.
Автогамия осуществляется разными способами: при прямом соприкосновении рыльца пыльников (контактная автогамия), при высыпании пыльцы из пыльников и оседании ее на рыльце под действием собственной тяжести (гравитационная автогамия), при помощи дождя, (омброавтогамия), ветра (анемоавтогамня), мельчайших насекомых, обитающих в цветке (трипсавтогамия).
Контактная автогамия наиболее обычна. В начале цветения, когда еще не утрачены шансы на перекрестное опыление, пыльники и рыльца в цветке созревают в разное время или расположены таким образом, что непосредственный контакт между ними невозможен. Позднее в цветке происходят изменения во взаимном расположении пыльников и рыльца. Они связаны с ростовыми движениями и проявляются в удлинении или изгибе тычинок или столбиков, благодаря чему вскрывшиеся пыльники и ставшие восприимчивыми рыльца располагаются на одном уровне и в непосредственной близости.
Контактную автогамию можно наблюдать у характерных растений темнохвойного леса (седмичника европейского, цирцеи альпийской, майника двулистного). У седмичника цветки слабо прототипичны. Рыльце восприимчиво к моменту распускания цветка. В это время пыльники находятся примерно на одном уровне с рыльцем и рядом с ним, что делает возможным их соприкосновение. Липкое рыльце прочно удерживает пыльник. Пыльник при покрывании высыпает на него пыльцу. Подобная же контактная автогамия встречается у цирцеи альпийской и майника двулистного. У последнего возможна также гейтоногамия между соседними цветками соцветия. У рассмотренных растений темнохвойного леса автогамия имеет более или менее случайный характер.Никаких структурных приспособлений к автогамии и связанных с ней редукций в цветке нет. Цветки этих растений вполне сохранили энтомофильный облик, но опыление насекомыми у них утрачено, вследствие чего они перешли к автогамин.
Копытень европейский — обычное растение в еловых лесах южной тайги и в смешанных и широколиственных лесах. Цветет в мае и первой половине июня. Цветки одиночные, невзрачные, располагаются у поверхности почвы, скрыты листьями. Венчик колокольчатый, спайнолепестный, снаружи буроватый, изнутри грязпо-пурпуровый. Долгое время способ опыления у копытня оставался неясным. Сейчас единодушно признается наличие у него обязательной контактной автогамии.
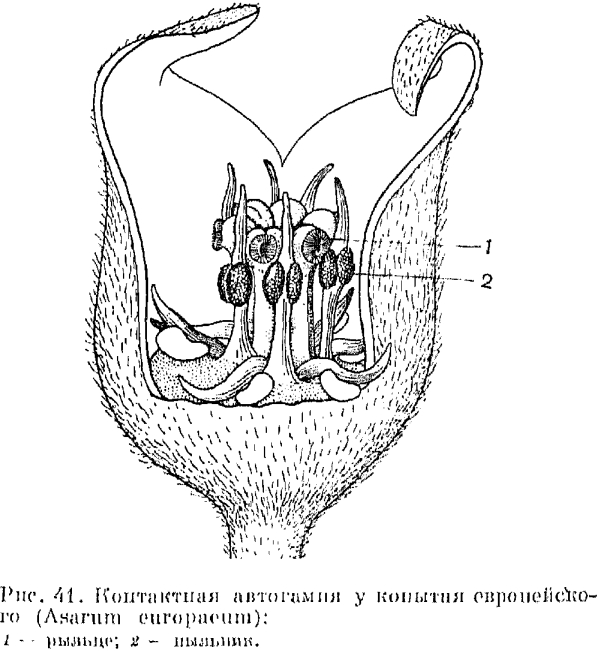
В развитии цветка копытня можно выделить три стадии —- начальную женскую, среднюю обоеполую и конечную мужскую, Для опыления имеет значение только средняя стадия. Цветки прототипичны: в бутоне и тотчас после распускания цветка рыльце восприимчиво, но пыльники 12 тычинок еще не вскрыты. В следующей, обоеполой стадии, при еще функционирующем рыльце, пыльники 6 длинных тычинок внутреннего круга вдвигаются между лопастями рыльца и, вскрываясь, оставляют на них пыльцу (рис. 41) на этой стадии легко можно обнаружить целые связки пыльцевых трубочек, возникших из пыльцевых зерен, находящихся еще во вскрытых пыльниках и врастающих в рыльце. Наконец, в последней, мужской стадии развития цветка рыльце уже утратило восприимчивость, а пыльники 6 наружных тычинок вскрываются, но, будучи более короткими, не достигают уровня рыльца. Поэтому эта конечная стадия, как и первая, не имеет сейчас значения для опыления, но могла иметь его в филогенетическом прошлом, когда копытень был еще энтомофильным растением. Облигатная (обязательная) автогамия копытня очень близка к клейстогамии. Иногда у него обнаруживаются закрытые цветки, в которых произошла автогамия.
С гравитационной автогамией лучше всего можно познакомиться на примере одноцветки (табл. 1.). Это очень красивое растение еловых лесов. Крупные белые венчики его как звездочки мерцают в сумраке леса. Они источают нежный аромат, но не посещаются насекомыми. Для осуществления автогамии у одноцветки необходима определенная ориентация цветоножки. Тогда, как это показано на рисунке 42, пыльца, высыпающаяся из пыльников, попадает на рыльце. То же самое, по-видимому, происходит в поникших цветках вересковых (брусинки, черники, голубики, подбела и грушанок круглолистной и средней, тогда как у грушанки малой имеется, видимо, контактная автогамия, поскольку пыльники у нее очень приближены к рыльцу). Ветер может содействовать гравитационной автогамии, поскольку при раскачивании и сотрясении растений происходит высыпание пыльцы через отверстия (поры), имеющиеся в пыльниках вересковых.
Своеобразный способ автогамии цветков при помощи дождя описан известным датским ботаником Олафом Хагерупом на Фарерских островах. Венчик цветка во время дождя наполняется до определенного уровня водой, причем пыльца плавает на ее поверхности. Пыльцевые зерна переносятся при помощи дождевых капель из пыльников на рыльце того же цветка. Насколько действен этот способ опыления, пока еще неизвестно, и по этому поводу высказываются сомнения.
Автогамии могут способствовать мельчайшио насекомые, обитающие в цветке (трипсы). это так называемая трипсоавтогамия. Она встречается у самых различных растений.
Клейстогамия представляет крайнюю форму автогамии. Она осуществляется в закрытых цветках, имеет экологический характер и вызывается неблагоприятными условиями внешней среды. Причины, вызывающие клейстогамию, разнообразны и у разных видов неодинаковы. Среди таких причин нужно назвать засуху, высокую атмосферную влажность, низкую или высокую температуру, условия освещения (глубокая тень, несоответствие длины фотопериода), затопление водой, скудное минеральное питание, неблагоприятные почвенные условия и т. д.
Клейстогамия выражена в разной степени. В соответствии с этим можно различать обязательную и факультативную клейстогамию. Под обязательной клейстогамией мы понимаем такие случаи, когда у вида постоянно образуются клейстогамные цветки, обнаруживающие черты более или менее далеко зашедшей редукции. Среди обязательных клейстогамов весьма обычны такие, у которых на одной и той же особи образуются нормальные хазмогамные и редуцированные клейстогамные цветки, причем именно последние являются плодущими. Развиваются эти разные формы цветков в разные сроки вегетационного периода, роже одновременно.
Клейстогамные цветки очень мелкие, никогда не раскрываются, напоминают бутоны. Чашечка уменьшена в размерах, лепестки рудиментарны или совсем отсутствуют, число тычинок часто уменьшено, пыльники мелкие, содержат мало пыльцы, которая не высыпается на рыльце, а прорастает в гнездах пыльника. При этом пыльцевые трубки прободают стенку пыльников и достигают рыльца. Последнее слабо развито. Эти цветки не выделяют нектар и не испускают аромат. В описанном случае развитие клейстогамии зашло далеко. Клейстогамия вызвала упрощение структуры цветка —- известную редукцию его органов и утрату ряда приспособлений к перекрестному опылению. Вместе с тем клейстогамия представляет важное приспособление к неблагоприятным условиям среды, обеспечивая постоянно высокую семенную продуктивность.
Хорошим примером такой постоянной, далеко зашедшей клейстогамии, сопровождающейся явлениями редукции в цветке, могут служить многие фиалки и кислица обыкновенная.
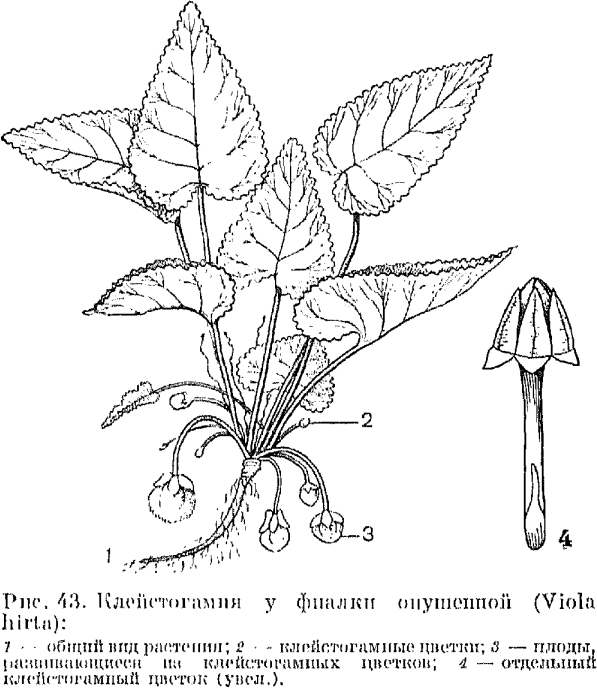
Фиалка удивительная — довольно обычное растение широколиственных и смешанных лесов. Она образует весной более или менее одновременно клейстогамные и хазмогамные открытоцветущие цветки. Последние в большинстве стерильны. У фиалки опушенной, произрастающей в широколиственных лесах, по сухим склонам,кустарниковым зарослям, ранней весной появляются на длинных цветоножках красивые цветки с крупными фиолетовыми венчиками. Они имеют нормально развитые тычинки и завязи, но, как правило, бесплодны. После их увядания на коротких цветоносах развиваются крохотные цветки; лепестки их прикрыты чашечкой. К моменту, когда начинает развиваться завязь, тычинки и лепестки в виде шапочки располагаются на рыльце. Из этих клейстогамных цветков образуются крупные многосемянные коробочки. Они сидят на удлинившихся к этому времени цветоносах, протягивающихся по поверхности земли (рис. 43). Клейстогамные цветки развиваются и у других фиалок (душистой, холмовой, собачьей).
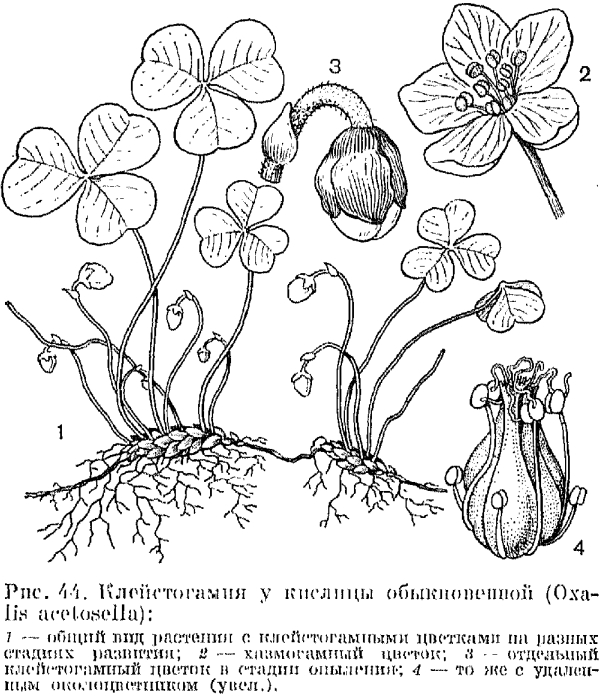
Кислица обыкновенная — характерное растение темнохвойной тайги. Она цветет в конце весны — начале лета (май — июнь). Крупные бело-розовые цветки ее многочисленны и являются украшением сумрачных еловых лесов. Из этих хазмогамных цветков образуется некоторое количество плодов-коробочек. Клейстогамные цветки кислицы мелкие, напоминают бутоны, не превышают 3 мм, сидят на коротких цветоножках, скрытых в моховом покрове (рис. 44). Появление их совпадает с началом созревания семян: в коробочках хазмогамных цветков (примерно спустя месяц после их цветения). В клейстогамных цветках кислицы лепестки сильно редуцированы и имеют вид крошечных чешуек, пыльники никогда не вскрываются, пыльца прорастает в пыльниках, пыльцевые трубки проникают через стенку пыльника и растут в сторону рыльца, нередко круто изгибаясь. По сравнению с хазмогамными цветками гинецей здесь отличается тем, что столбики у него короче и сосочки на рыльцах развиты слабее, а иногда столбиков совсем нет и рыльцевая поверхность располагается на вершине плодолистиков. Клейстогамия у кислицы обусловлена сильным затенениоми отчасти повышенным или избыточным увлажнением.
Особую группу составляют растения с подземными клейстогамными цветками. Лучше других известен из этой группы арахис подземный, называемый также китайскими орешками. Это культурное масличное растение родом из Южной Америки. У нас его возделывают на юге Украины, Северном Кавказе, в Закавказье и Средней Азии. У арахиса бобы образуются в почве. С самого начала они развиваются под землей из подземных клейстогамных цветков. Однако у арахиса имеются еще и надземные цветки с оранжево-желтыми венчиками, расположенные у самой поверхности почвы или несколько выше на стебле. После опыления этих цветков завязи их внедряются в почву на глубину 8 см вследствие удлинения ножки (гинофора), на которой сидит завязь.
Факультативная клейстогамия, будучи по своей природе также экологической, представляет как бы начальную фазу развития данного явления — она далеко не столь постоянна, проявляется лишь временами, при определенном сочетании условий среды и не сопровождается (это самое главное) признаками редукции в цветке. Факультативная клейстогамия довольно обычна. Примером ее могут служить ковыли, хлебные и другие злаки, отдельные виды пустынных маревых, некоторые водные и болотные растения — частуха подорожниковая, вахта трехлистная, виды рода росянка (Droseга), вербейник обыкновенный и др.
Превосходный пример факультативной клейстогамии представляют ковыли — характернейшие растения степей. У ковылей бывает открытое (хазмогамное) и закрытое (клейстогамное) цветение. Открытое цветение и перекрестное опыление при помощи ветра происходит у них ночью и на рассвете, но бывает далеко не всегда. Нередко ковыли полностью или частично цветут клейстогамно. Клейстогамия вызывается у них почвенной засухой (ксероклейстогамия) или пониженной температурой (термоклейстогамия). Клейстогамия у ковылей усиливается, и становится преобладающей в болое сухие сезоны и в более засушливых районах.
Экологический характер имеет клейстогамия у хлебных злаков. У овса она вызывается высокой или очень низкой температурой, а у ячменя — недостатком тепла и дождливой погодой на севере и засухой в степной золе. Что касается пшениц, то представления об условиях, способствующих или препятствующих открытому цветению их, очень противоречивы. Пo-видимому, теплая и влажная погода способствует открытому цветению, тогда как засуха и высокая температура — клейстогамному.
Итак, самоопыление у цветковых растений представляет явление вторичное, вызванное неблагоприятными для ксеногамного опыления условиями среды. Соотношение самоопыления и разных способов перекрестного опыления зависит от экологических условий. Эта зависимость отчетливо выступает в темнохвойной тайге, Арктике и пустынях, поскольку именно здесь крайние условия часто препятствуют осуществлению перекрестного опыления.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
В случае обоеполости цветков наблюдаются: 1) автостерильность, или самобесплодность — неспособность пыльцы прорастать на рыльце пестика того же самого цветка (встречается у большинства сортов яблони, груши, вишни, некоторых орхидных); 2) дихогамия — неодновременное созревание в обоеполых цветках тычинок и пестиков; она выражена в двух формах: протероандрия и протерогиния; при протероандрии пыльники созревают раньше пестиков (виды семейств бобовых, гвоздичных, гераниевых, астро вых, зонтичных); при протерогинии пестики в цветках созревают раньше тычинок (капустные, розовые, злаки); 3) такое расположение тычинок и пестиков, при котором пыльца не может попасть на рыльце пестика того же самого цветка (вероника, губоцветные); 4)гетеростилия, или разностолбчатость, когда у одних особей в цветках образуются длинные столбики, превышающие длину тычинок, у других в цветках столбики короткие, а тычинки — длинные (примула, медуница)
Самоопыление имеет одно существенное преимущество по сравнению с перекрестным: оно не зависит от погодных условий и посредников, поэтому осуществляется при любых условиях.
Дихогамия — это функциональная раздельнополость. Она вызвана разновременным созреванием и экспонированием пыльцы и рыльца в цветках, вследствие чего последние выступают то в мужской (пыльниковой), то в женской (рыльцевой) фазе, Дихогамия проявляется в форме протандрии (при более раннем созревании пыльцы) или протогинии (при более раннем созревании рылец). Одновременное созревание пыльцы и рылец в цветке носит название гомогамии.
Таблица 1. Приспособления цветков к опылению:
1 - протандрия у кипрея узколистного, или иван-чая (Chamaenerium angustifolium); 2 - пыльценосный цветок у зверобоя продырявленного (Hypericum perforatum); 3 - взрывчатое цветение у костреца безостого (Bromopsis inermis); 4 - гравитационная автогамия у одноцветки крупноцветковой (Moneses uniflora); 5 - съедобные волоски на тычинках у коровяка медвежьего уха (Verbascum tapsus); 6 - соцветие с наружными бесполыми цветками у калины обыкновенной (Viburnum opulus).


Рис. 31. Примеры протандрии.
Скабиоза бледно-желтая (Scabiosa ochroleuca): 1 - краевой цветок в тычиночной стадии; 2 - он же в рыльцевой стадии; 3 - срединный цветок в тычиночной стадии; 4 - он же в рыльцевой стадии. Смолевка вильчатая (Silene dichotoma): 5 - цветок в тычиночной стадии первого дня цветения, 6 - он же в тычиночной стадии второго дня цветения, 7 - он же в рыльцевой стадии третьего дня цветения (все тычинки увяли).
Дихогамия обычно рассматривается как приспособление к перекрестному опылению и как средство предотвращения самоопыления. Это традиционное представление о дихогамии нуждается в уточнении. Дихогамия необязательно исключает самоопыление. Нередко в конце цветения возможна автогамия, если по каким-либо причинам перекрестное опыление не осуществилось. Кроме того, самоопыление в форме гейтоногамии возможно у большинства дихогамных растений еще и потому, что на них имеются многочисленные цветки, находящиеся в разных фазах развития. Поэтому значение дихогамии следует усматривать в том, что она более или менее ограничивает самоопыление, способствуя тем самым перекрестному опылению. Такое сочетание перекрестного опыления и самоопыления играет, как уже говорилось, положительную роль в эволюции.
Протандрия встречается чаще, чем протогиния. Она более соответствует нормальной последовательности в развитии частей цветка. Протандрия хорошо выражена у зонтичных, сложноцветных, колокольчиковых, гвоздичных, ворсянковых, губоцветных и многих других семейств (рис. 31, табл. 1).
У колокольчиков пыльники вскрываются еще в бутоне. Пыльца муфточкой окружает столбик, удерживаясь на волосках, покрывающих его. Опорожненные и подсохшие пыльники видны на дне венчика. Лопасти рыльца в этот момент еще вполне сомкнуты. Примерно сутки спустя после распускания цветка расходятся лопасти рыльца и становится возможным опыление их чужой пыльцой, принесенной насекомыми. Но под конец цветения возможна и автогамия, благодаря тому что лопасти рыльца, закручиваясь спирально вниз, касаются воспринимающей поверхностью столбика, сохранившего собственную пыльцу.

Рис. 32. Протогиния у подорожника Корнута (Plantago cornutii):
1 - цветок в стадии бутона; 2 - появление рыльца; 3 - увядание рыльца; 4 - раскрывание цветка и начало выдвижения тычинок при уже увядшем рыльце; 5 - тычиночная стадия цветка.
У зонтичных и ворсянковых протандрия является весьма действенным средством предотвращения самоопыления.
У зонтичных протандрия строга, безупречна и охватывает не только отдельные сложные зонтики, но и всю особь в целом. У большинства видов зонтичных это достигается благодаря строгой очередности в цветении зонтиков разного порядка и полной согласованности (синхронности) его у зонтиков данного порядка. Вследствие этого каждая особь последовательно несколько раз выступает то в пыльниковой, то в рыльцевой фазе, причем эти фазы резко разграничены и, как правило, не налегают друг на друга. В качестве примера может служить порезник средний (Libanotis intermedia). Этот тип протандрии (тип Libanotis) с многократной сменой тычиночной и рыльцевой фаз очень обычен у зонтичных. Значительно реже у них встречается другой тип протандрии (тип Peucedanum), характеризующийся однократной сменой фаз у всех одновременно и согласованно цветущих зонтиков особи, независимо от того, к какому порядку они принадлежат. Такая протандрия известна пока у горичника Любименко (Peucedanum lubimenkoanum).
Резкая протандрия, всего соцветия, последовательная и одновременная смена фаз в цветении соцветий разных порядков свойственна и некоторым ворсянковым: короставнику полевому (Knautia arvensis), скабиозе бледно-желтой (Scabiosa ochroleuca) и др. У зонтичных и ворсянковых в результате этого невозможна автогамия и гейтоногамия, неизбежным является перекрестное опыление.

Рис. 33. Протогиния у ситника Жерарда (Juncus gerardii):
1 - цветок в рыльцевой фазе вечером (протогиния); 2 - раскрывшийся цветок утром в момент опыления.
Протогиния хорошо выражена у крестоцветных, розовых, барбарисовых, жимолостных и т. д. Во многих случаях разница во времени созревания рыльца и пыльцы столь незначительна, что наличие протогинии кажется сомнительным. Более резко выражена протогиния у ветроопыляемых растений, причем не только у обоеполых, но и у однодомных и двудомных растений. Так обстоит дело у ситниковых, осоковых, злаковых, полыней, подорожников (рис. 32). У ситников популяция вида вступает в рыльцевую фазу с вечера предшествующего дня, а на следующее утро переходит в тычиночную фазу, причем рассеивание пыльцы ограничено 2-3 ч (рис. 33). У однодомных осок рыльцевая фаза предваряет пыльниковую на 1-6, а у подорожников на 4-6 суток.
В подобных случаях роль протогинии заключается в том, что заблаговременная экспозиция рылец представляет подготовку к очень быстротечному опылению ветром во время кратких суточных периодов рассеивания пыльцы. Ту же функцию выполняет протогиния у некоторых пустынных маревых.

Рис. 34. Гетеростилия у примулы (Primula sp.):
1-4 - короткостолбчатая форма (1 - вид цветка в разрезе; 2 - короткий столбик с сосочковидной поверхностью рыльца при малом увеличении; 3 - то же при большом увеличении; 4 - пыльцевое зерно); 5-8 - длинностолбчатая форма (5 - вид цветка в разрезе; 6 - длинный столбик с сосочковидной поверхностью рыльца при малом увеличении; 7 - то же при большом увеличении; 8 - пыльцевое зерно).
Явление гетеростилии состоит в том, что у вида имеются две или три формы цветков, находящиеся на разных особях и различающиеся по длине столбиков и расположению тычинок (диморфные и триморфные растения). У диморфных первоцветов (Primula) длинностолбчатая форма имеет цветки с длинным столбиком, рыльце которого находится в зеве венчика, а тычинки — ниже, в глубине венчика. У короткостолбчатой формы обратное расположение столбиков и пыльников (рис. 34). Формы гетеростильных растений различаются и по другим признакам. Так, короткостолбчатые цветки характеризуются по сравнению с длинностолбчатыми более крупной пыльцой и более мелкими сосочками рыльца. Самоопыление у той и другой формы, а также переопыление между особями одного морфологического типа дает ничтожное количество семян (самонесовместимость), тогда как переопыление между растениями с разной длиной столбика высокоэффективно. Примером диморфных (дистильных) видов могут служить медуница, первоцвет, гречиха, кермек и др.
У триморфного гетеростильного дербенника иволистного (Lythrum salicaria) имеются коротко-, средне- и длинностолбчатые цветки, распределенные по разным особям. Тычинки в цветке каждой формы соответствуют по длине столбикам цветков двух других форм. Оплодотворение оказывается наиболее эффективным при опылении рыльца каждой формы цветка пыльцой тычинок соответственной длины из двух других форм цветка, как это было показано еще Ч. Дарвином (рис. 35).
Гетероморфная самонесовместимость встречается редко. Она известна у 56 родов из 23 семейств. Особенно она распространена у мареновых (Rubiaceae), среди которых насчитывается 155 гетеростильных видов.

Рис. 35. Гетеростилия у дербенника иволистного (Lythrum salicaria): 1 - длинностолбчатая форма; 2 - среднестолбчатая форма; 3 - короткостолбчатая форма.
Самоопыление растений
Перенос пыльцы с тычинок на рыльце того цветка называется самоопылением. У самоопылителей и тычинки, и пестики на одном цветке созревают одновременно. У растений в ходе длительного эволюционного развития выработались приспособления для перекрестного опыления, т.к. оно является прогрессивным. Образуется потомство с признаками материнского и отцовского организмов, а при самоопылении у потомства проявляются признаки лишь одного организма.
Самоопыление ведет к снижению урожайности, к вырождению сорта. Поэтому наиболее распространено перекрестное опыление и цветки большинства растений имеют приспособления, препятствующие самоопылению:
1. однополость (орешник, конопля, осина и др.);
2. неодновременное созревание пыльников и рылец;
3. различная длина тычинок и пестиков.
Однако самоопыление имеет значение в селекции при выведении чистых линий и может быть запасным актом на тот случай, если не произойдет перекрестного опыления.
Самоопыление в естественных условиях встречается реже, чем перекрестное опыление.
В естественных условиях не все цветки могут опылиться. Часто пыльца теряет свои свойства из-за воздействия слишком высоких температур или повышенной влажности. В безветренную погоду или, наоборот, при шквальном ветре опыление ветроопыляемых растений трудновыполнимо.
Из культурных растений размножаются путем самоопыления, например, пшеница, вика, соя, горох, ячмень и др. Такие растения как гречиха, клевер красный, эспарцет, яблони большинства сортов, нуждаются только в перекрестном опылении. При этом следует отметить, что перекрестное опыление способствует получению более мощных, плодовитых и жизнеспособных растений. Есть группа растений, которые сохранили способность к самоопылению (подсолнечник, люцерна, хлопчатник, малина, крыжовник и некоторые другие), но и они при перекрестном опылении насекомыми дают более высокие урожаи.
У многих растений в процессе длительного эволюционного развития выработался целый ряд приспособлений, препятствующих самоопылению и способствующих перекрестному опылению. У целой группы растений невозможно самоопыление, потому что у них женские цветки с пестиками развиваются на одном экземпляре, а мужские с тычинками на другом (однополые). Такие растения принято называть двудомными. В эту группу входят ива, тополь, конопля, клубника и др. Есть растения, у которых цветки, так же как и у двудомных растений, имеют или тычинки или пестики, но они находятся на одном и том же экземпляре. Такие растения называют однодомными.
Препятствием самоопылению является неодновременное созревание тычинок и пестиков, находящихся в одном и том же цветке (обоеполый цветок). Так, у подсолнечника, крыжовника, кипрея пыльники созревают раньше, чем рыльце, а у яблони, груши, подорожника и других раньше созревает рыльце. В этих случаях опыление происходит пыльцой с других цветков, перенесенной насекомыми. У ряда растений (гречиха, медуница, дербенник) самоопыление затруднено из-за того, что цветки имеют разностолбчатые органы: у одних длинные тычинки и короткие пестики, у других наоборот — длинные пестики и короткие тычинки. Поэтому в пределах цветка опыление собственной пыльцой, как правило, не осуществляется.
У некоторых растений (клевер красный, эспарцет) наблюдаются явления самобесплодности (самостерильности). В этом случае собственная пыльца на рыльце цветка не прорастает или прорастает значительно медленнее, чем попавшая с другого цветка. Самобесплодность встречается у яблони, груши, черешни и ряда других плодовых и ягодных культур, оплодотворение у которых происходит лишь в том случае, если на рыльце пестиков одного сорта попала пыльца с цветков другого сорта. Чтобы создать условия для нормального плодоношения этих культур, необходимо в садах иметь соответствующие сорта плодовых деревьев, обеспечивающие межсортовое опыление.
У множества цветковых растений выработан механизм невозможности самоопыления, что предотвращает родственное скрещивание и увеличивают генетическое разнообразие вида. Это достигается благодаря процессу, названному самонесовместимостью, при котором опыление не в состоянии произойти, если пыльца идентифицирует попадание на пестик цветка, на растении которого она появилась. Но принцип действия этого механизма долго оставался загадкой, которая теперь успешно решена.
Группа исследователей во главе с Те-Хуи Као, профессора биохимии и молекулярной биологии в Пенсильванском Университете, сообщает в статье, опубликованной 20 мая в выпуске журнала Nature, об открытии у петуний гена, который контролирует этот процесс самонесовместимости пыльцы. Недавно идентифицированный ген получил название PiSLF (от Petunia inflata S-locus F-box). Этот ген кодирует производство большого семейства белков F-box, которые, как известно, вызывают сворачивание белка в различных живых организмах, включая животных, растения и дрожжи.
В то время как разновидность петунии может нести от 50 до 60 различных S-аллелей, каждое растение имеет только две из них, по одной от каждого родителя. Аллель - один из множества возможных вариантов специфического гена; например, две аллели существуют для каждого из трех генов, которые определяют цвет глаза у людей. Зерно пыльцы является гаплоидным. Это означает, что оно содержит только половинный набор хромосом, и таким образом каждое зерно пыльцы содержит только одну из двух S-аллелей родительского растения. Пестик, с другой стороны, является диплоидным, и имеет два набора хромосом (по одному от каждого родителя) и поэтому имеет обе S-аллели родительского растения. Во время опыления, если S-аллель пыльцы не соответствует любой из этих двух S-аллелей в пестике, то пыльца прорастет через поверхность рыльца пестика. Из ее вегетативной клетки развивается пыльцевая трубка, которая несет к зародышевому мешку спермии. Достигнув его, спермии сливаются с яйцеклеткой, и происходит оплодотворение. Однако, если S-аллель пыльцы соответствует любой из этих двух S-аллелей в пестике, рост пыльцевой трубки останавливается на одну треть пути к мешку, что предотвращает оплодотворение. Вызов этого ответа самонесовместимости требует взаимодействия между генами S-аллели пыльцы и пестика. Ответственным за эту несовместимость оказался ген PiSLF.
Таким образом, открываются большие возможности генной инженерии в плане остановки безудержного скрещивания трансгенных растений с дикими. Ученым уже удалось переделать трансгенные растения в самоопыляемые (то есть те становятся самосовместимыми), и все потомство трансгенных растений также наследует эту совместимость.
Это открытие может также получить коммерческое применение для гибридного производства злаковых культур. Развитие гибридных сортов является главной целью садоводческой и сельскохозяйственной практики, потому что гибридные растения наиболее производительные, чем сортовые культуры, полученные из самоопыления или случайного опыления. Однако, чтобы получить гибридный сорт, необходимо перекрестное опыление с двух определенных сортов, при котором самоопыление и родственное опыление (опыление растением того же самого сорта) необходимо исключить. До сих пор для этого применяли ручной метод опыления. Однако этот процесс является очень трудоемким и дорогим. Из-за этого становятся очень дорогими и семена гибридных сортов, а, следовательно, и будущая цена урожая и продуктов питания на нашем столе. Если удастся сократить затраты за счет снижения ручного труда при производстве гибридов, это значительно снизит цены на продукты питания.
Читайте также: