
Спорогенез и гаметогенез у цветковых растений и процесс оплодотворения
Обновлено: 04.10.2024
Особенности гаметогенеза у растений. Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор; 2-й этап — гаметогенез — включает образование зрелых гамет.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) —мега- или макроспорогенезом.
Микроспорогенез протекает аналогично сперматогенезу у животных, вплоть до образования сперматид, а мегаспорогенез соответствует образованию ооцита П.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У растений отсутствует зачатковый путь в том смысле, как это имеет место у животных.
Микроспорогенез и микрогаметогенез. Рассмотрим микроспорогенез и микрогаметогенез на наиболее общем примере покрытосеменных растений. В субэпидермальной ткани молодого пыльника, называемой археспорием, каждая клетка после ряда делений становится материнской клеткой пыльцы, которая проходит все фазы мейоза.
52
В результате двух мейотических делений возникают четыре гаплоидные микроспоры (рис. 24, /). Они лежат четверками, их называют тетрадами спор.
При созревании тетрады распадаются на отдельные микроспоры. Этим заканчивается микроспорогенез.
Вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, не способны к движению и называются спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три клетки. Две из них — спермин и одна вегетативная.
Мегаспорогенез и мегагаметогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, часто только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры (см. рис._ 24, 2).
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады гаплоидна. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития). Судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти, и затем ее ядро претерпевает ряд митозов. При этом сама клетка не делится, она образует зародышевый мешок.
У разных растений число митозов может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) бывает три деления, в результате возникает восемь одинаковых ядер. Во время этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других в противоположном конце зародышевого мешка, называемого халазальным (рис. 25). В дальнейшем ядра обособляются в самостоятельные клетки, имеющие значительное количество цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка и две так называемые синергиды — обра-
53
зуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 одинаковых гаплоидных ядер, из которых только одно образует яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
Кроме моноспорического типа развития, который только что рассмотрен, существуют и другие. При различных типах развития зародышевого мешка сохраняется разное' 1 количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям (если сохраняются две мегаспоры — биспорический; четыре — тетраспорический тип развития).
Сравнение процессов созревания половых клеток у животных и растений (рис. 23 и 24) показывает почти полный параллелизм их, несмотря на то, что расхождение (дивергенция) растений и животных в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов в растительном и животном мире.
ОПЛОДОТВОРЕНИЕ
Сущность оплодотворения. Оплодотворением принято называть побуждение яйца к развитию путем объединения в нем ядер (кариогамии) мужских и женских половых клеток. Оплодотворение представляет собой необратимый процесс; однажды оплодотворенное яйцо не может быть оплодотворено вновь. Син-гамия (слияние мужских и женских половых клеток) и кариогамия составляют сущность процесса оплодотворения.
Оплодотворение у животных. Процесс оплодотворения у животных можно разделить на несколько фаз. Первая фаза начинается с того, что сперматозоид либо прикрепляется к любой точке поверхности яйцеклетки, либо проникает в нее через микропиле. Момент соприкосновения головки сперматозоида с яйцом является начальным в цепи химических реакций. Эту фазу называют фазой активации яйца.
К моменту соприкосновения сперматозоида с яйцом и проникновения его внутрь ядро яйца у разных животных может находиться на разных стадиях деления созревания. Кариогамия же может произойти только после окончания мейоза, т. е. после образования женского пронуклеуса.
В акте оплодотворения два гаплоидных пронуклеуса сливаются в одно ядро, образуя зиготу. Этот момент является кульминационным пунктом процесса полового размножения. В результате кариогамии гомологичные хромосомы, разошедшиеся в мейозе у предыдущего поколения, вновь воссоединяются в одном ядре зиготы. Так восстанавливается диплоидный набор хромосом при половом размножении.
В цитоплазму яйцеклетки у млекопитающих проникает не только головка (ядро) сперматозоида, как это считали раньше, но и его шейка и даже хвостовая часть, что создает возможность передачи некоторого количества цитоплазмы мужского организма потомству.
Оплодотворение у растений. Оплодотворение у растений в принципе сходно с таковым у животных, но имеет и некоторые особенности.
Раньше было сказано, что микрогаметогенез завершается образованием двух спермиев, которые формируются или в пыльцевом зерне, или в пыльцевой трубке при прорастании пыльцевого зерна.
Пыльцевая трубка, дорастая до микропиле зародышевого мешка, соприкасается с яйцевым аппаратом — яйцеклеткой и синергидами (рис. 26, /). При соприкосновении конца пыльцевой трубки с синергидами пыльцевая трубка лопается, а синергиды разрушаются. Передвигающиеся по пыльцевой трубке по мере ее роста два генеративных ядра — спермин после разрыва трубки вместе с ее содержимым попадают внутрь зародышевого мешка (рис. 26, 2). Из двух проникших в зародышевой мешок спермиев один спермин сливается с гаплоидным ядром яйцеклетки (рис. 26, 3). Слияние ядра спермия с ядром яйцеклетки является собственно оплодотворением у растений. В оплодотворенной яйцеклетке — зиготе восстанавливается диплоидное число хромосом. Из зиготы развивается зародыш семени.
Гаметогенезом называют процесс образования половых клеток (гамет). Этот процесс происходит у мужских и женских особей в гонадах (половых железах), представленных семенниками (яичками) и яичниками.
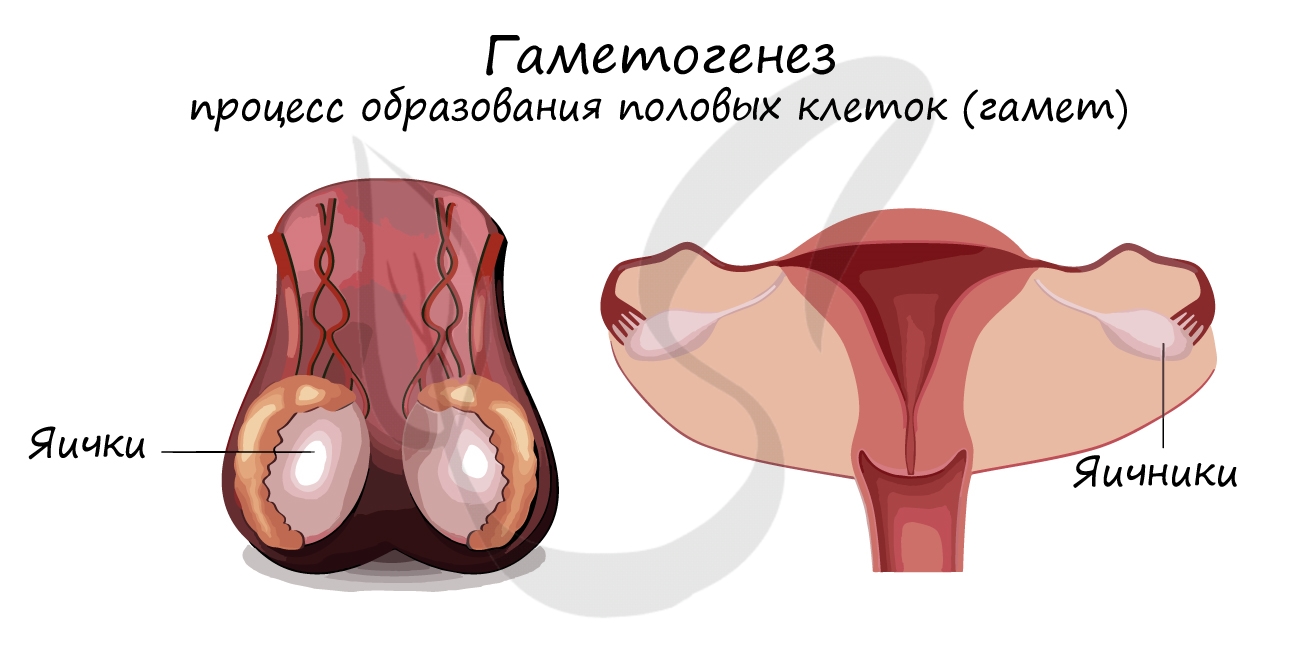
Гаметы (n) образуются в результате мейоза из клеток-предшественников (2n, как у соматических клеток). Половые клетки гаплоидны, то есть имеют в два раза меньшее число хромосом, чем клетки-предшественники. Мужская (n) и женская (n) гаметы, сливаясь друг с другом в процессе оплодотворения, образуют зиготу (2n).
Таким образом, за счет гаплоидности гамет (в результате мейоза) поддерживается постоянное количество хромосом в ряду поколений, не происходит их удвоения.
Процессы сперматогенеза и овогенеза (оогенеза) требуют нашего более детального изучения.
Сперматогенез (греч. sperma – семя + genesis – зарождение)
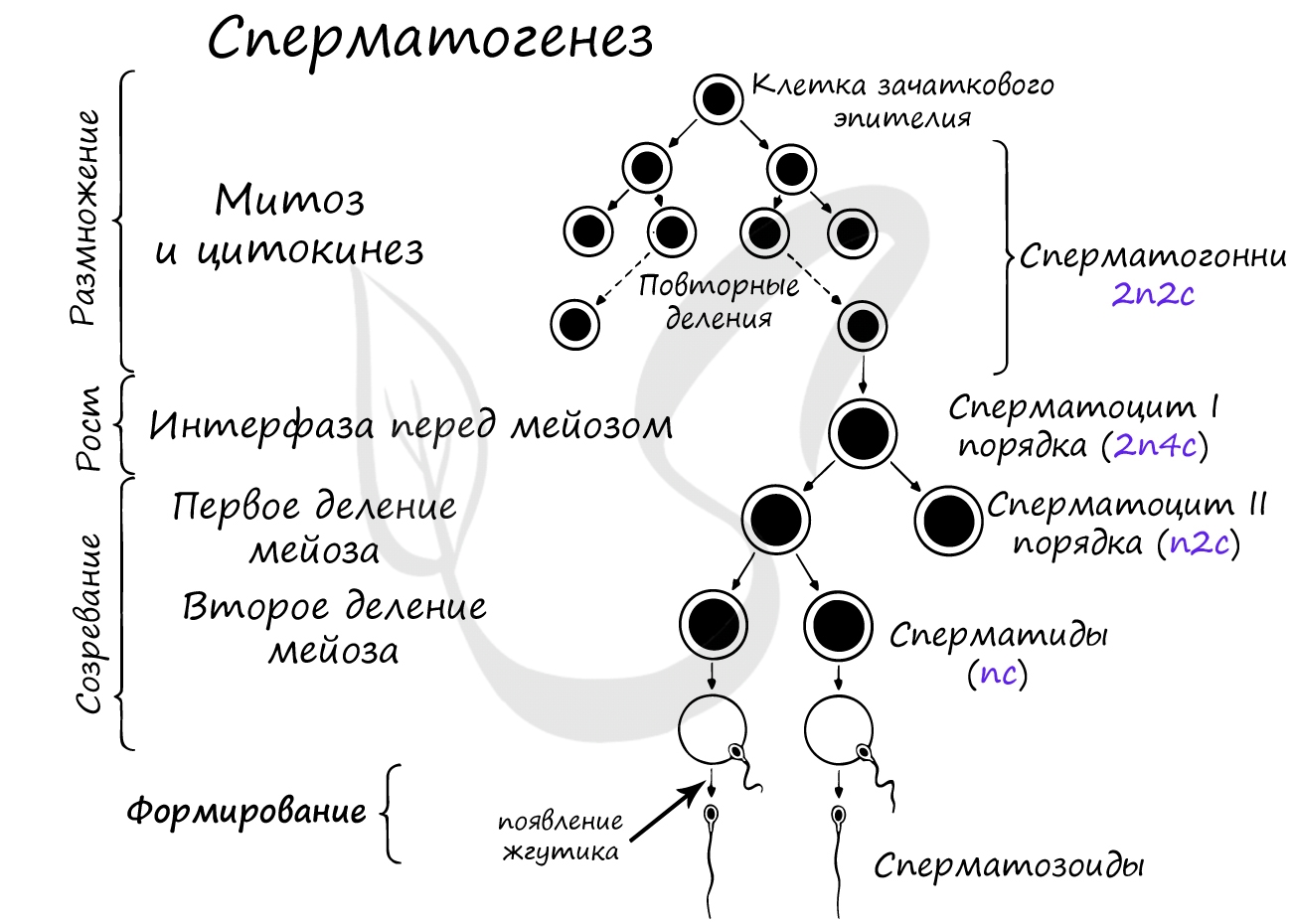
В ходе фазы размножения диплоидные сперматогенные клетки (2n2c) многократно делятся митозом, в результате образуются сперматогонии (2n2c) - стволовые клетки. Часть сперматогоний вступает в последующее митотическое деление, образуя такие же сперматогонии (2n2c).
Половые клетки в этой фазе называются сперматоцитами I порядка, они теряют способность к митотическому делению.
В этот период клетка растет, увеличивается количество органоидов и цитоплазмы. Происходит подготовка к мейозу, который начинается в следующей фазе - созревания.
На фазу роста приходится S-период: происходит удвоение ДНК, в результате чего набор хромосом сперматоцита I порядка становится (2n4c).
Происходит первое деление мейоза (мейоз I). В результате из сперматоцитов I порядка (2n4c) образуются сперматоциты II порядка (n2c). Между мейозом I и мейозом II практически отсутствует интерфаза, поэтому сперматоциты II порядка (n2c) сразу же вступают в мейоз II, в результате которого образуются сперматиды (nc).
Итак, в фазу созревания происходят первое и второе деления мейоза, которые приводят к тому, что образовавшаяся клетка - сперматида - имеет гаплоидный набор хромосом (nc).
В этой фазе у каждой сперматиды отрастает жгутик, после чего они получают полное право называться сперматозоидами. У основания жгутика концентрируются митохондрии - "энергетические станции клетки", которые всегда будут готовы предоставить АТФ для его активной работы.
Овогенез, или оогенез (греч. ōón — яйцо + genesis – зарождение)
Оогенезом называют процесс формирования женских гамет (половых клеток) - яйцеклеток. Он активируется в женском организме в период полового созревания (под действием женских половых гормонов) и длится до менопаузы (45-55 лет).
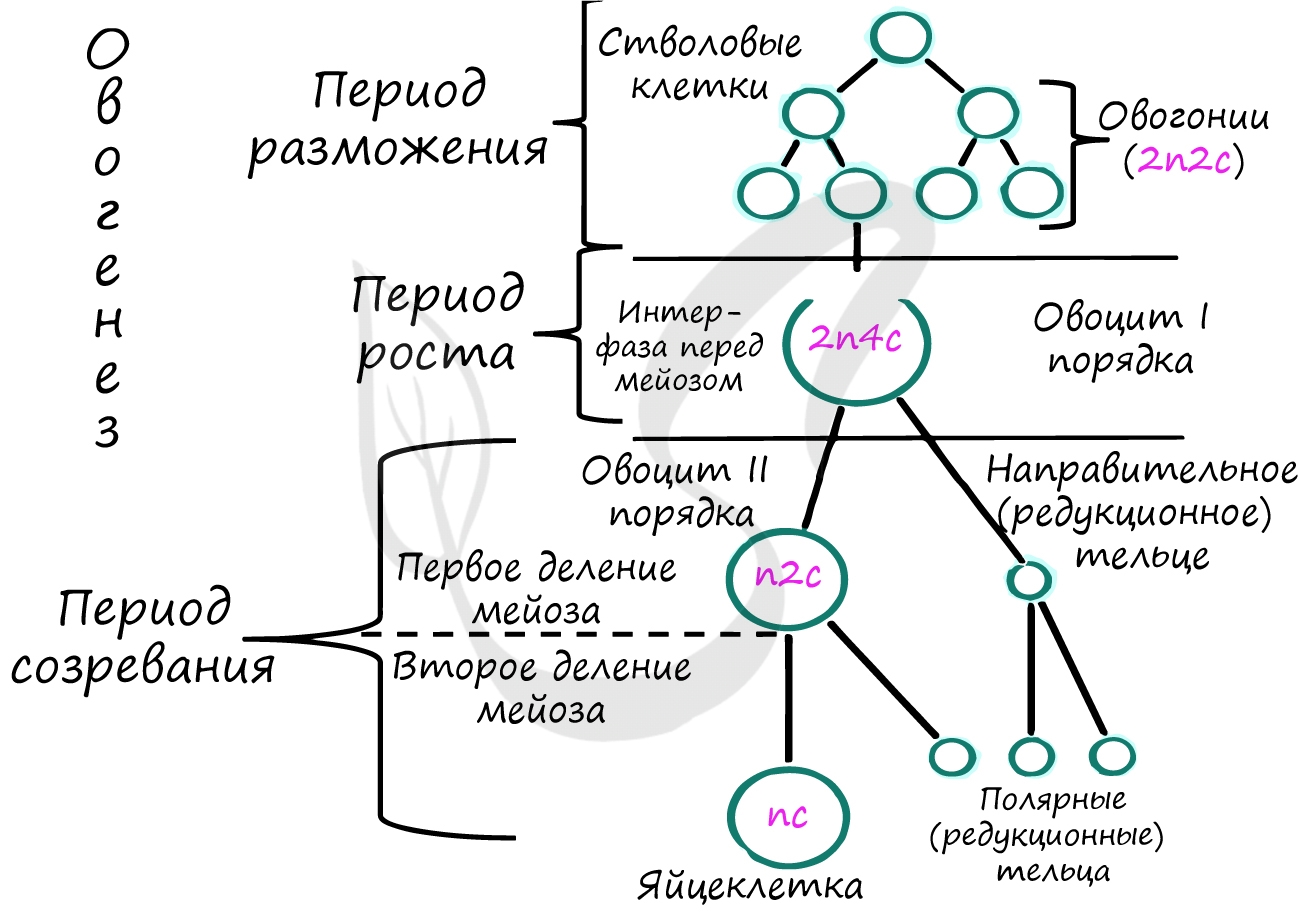
В результате многократных делений клеток яичника образуются стволовые клетки - овогонии (2n2c).
Половые клетки в этой фазе называются ооцитами I порядка, они теряют способность к митотическому делению.
В овогенезе эта фаза отличается более длительной продолжительностью, по сравнению с такой же фазой в сперматогенезе. Клетки накапливают большой запас питательных веществ. В этот период происходит удвоение ДНК в S-периоде - набор хромосом и ДНК ооцитов I порядка становится 2n4c.
Ооциты I порядка (2n4c) вступают в первое деление мейоза, в результате которого образуются ооциты II порядка (n2c) и первое полярное (направительное) тельце, которое не несет большой функциональной значимости и подвергается дегенерации.
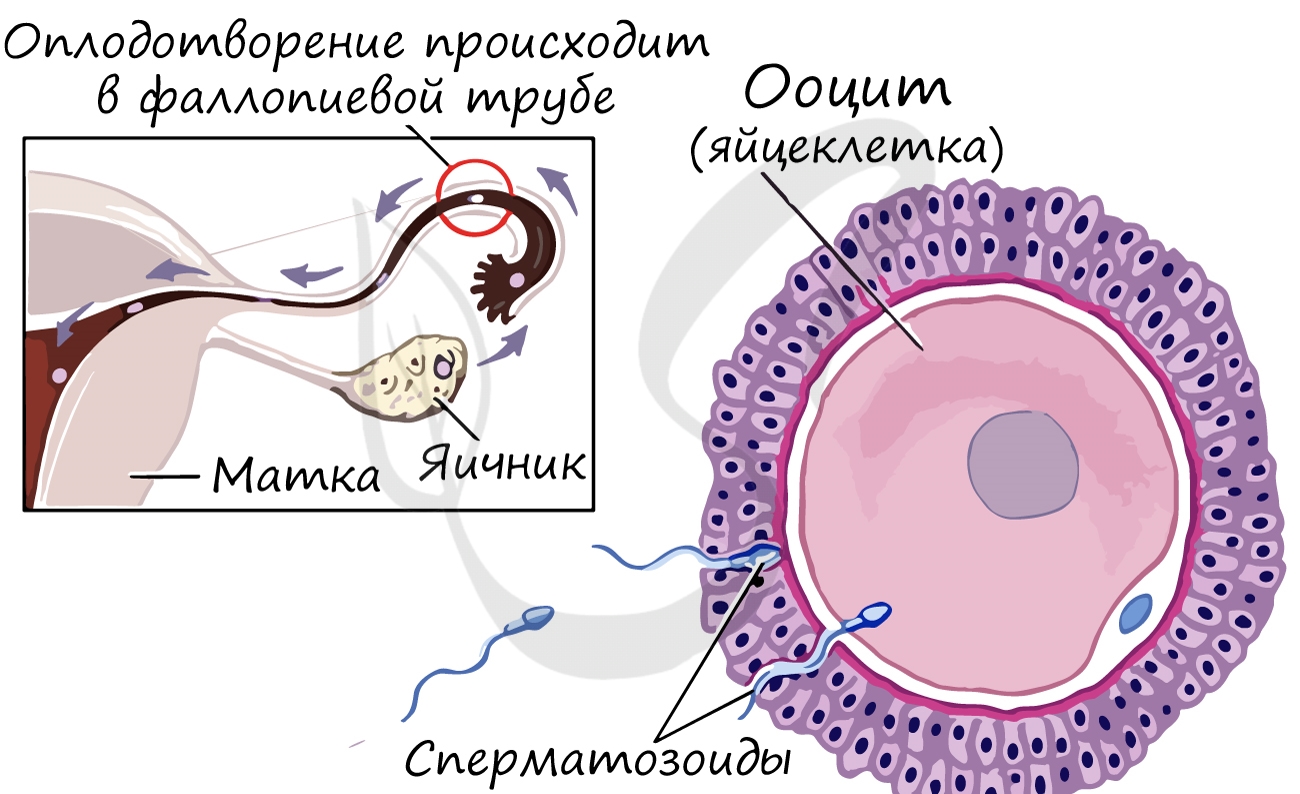
Второе деление мейоза начинается только после взаимодействия овоцита II порядка (n2c) со сперматозоидом. В результате этого образуется яйцеклетка (nc) и второе полярное тельце, которое также подвергается дегенерации.
Строго говоря, при овуляции из яичников выходит не "яйцеклетка", а ооцит II порядка, который ждет встречи со сперматозоидом для продолжения деления и развития будущего зародыша. Если такого взаимодействия не происходит, то яйцеклетка подвергается дегенерации.
Оплодотворение
Оплодотворение - ключевой процесс полового размножения, обусловленный слиянием сперматозоида и яйцеклетки. После оплодотворения в результате ряда стадий образуется эмбрион.
Сперматозоид (nc) обладает положительным химическим таксисом к яйцеклетке (nc). Оплодотворение - слияние сперматозоида с яйцеклеткой и образование зиготы (2n2c).
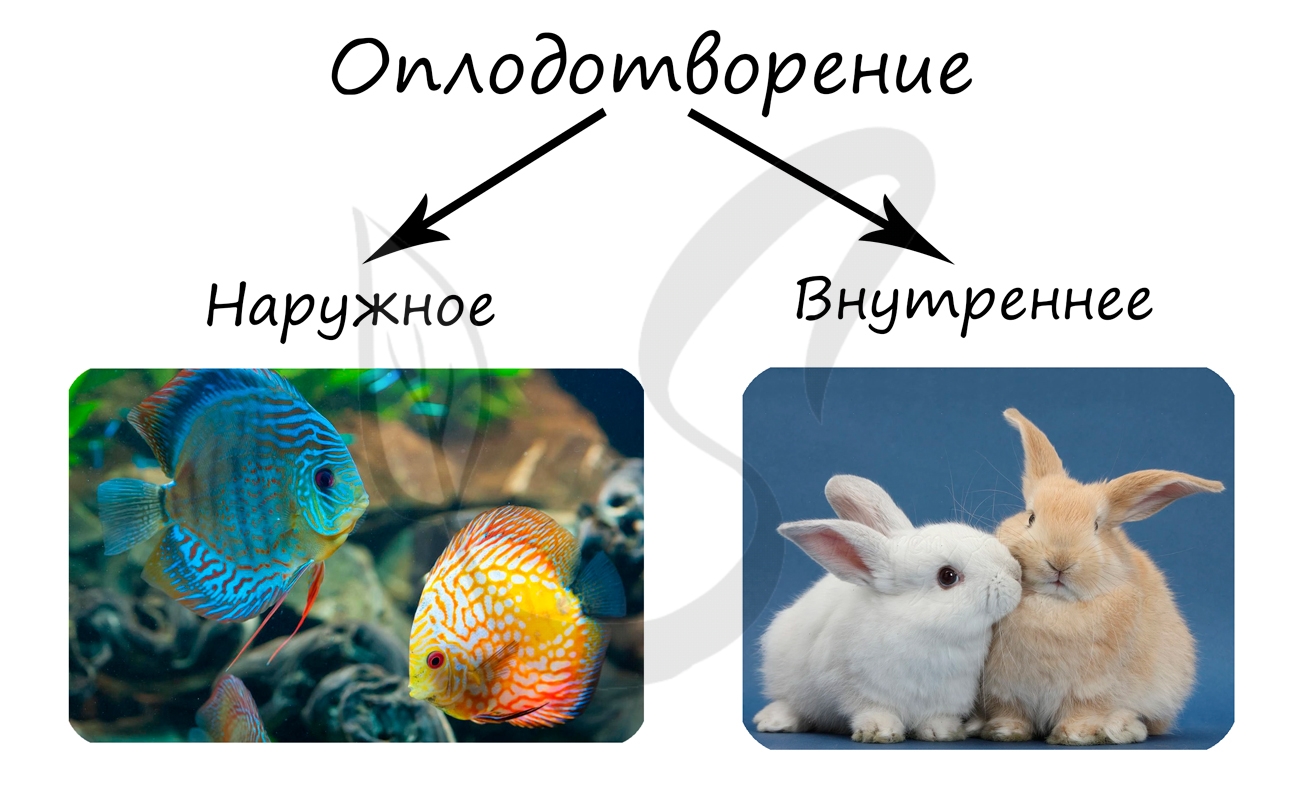
При внутреннем оплодотворении сперматозоид сливается с яйцеклеткой в женских половых путях, куда самец вводит семенную жидкость со сперматозоидами.
При внешнем оплодотворении сперматозоид сливается с яйцеклеткой вне половых путей самки, например, у двустворчатых моллюсков оплодотворение происходит в мантийной полости самки.
Внешнее оплодотворение характерно для рыб, земноводных, моллюсков. Внутреннее - для пресмыкающихся, птиц и млекопитающих.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Процесс развития и образования половых клеток называется гаметогенезом. Существенным этапом в этом процессе является мейоз, однако он не исчерпывает всего процесса гаметогенеза, который обладает спецификой у особей разных полов и у представителей разных групп организмов. Для понимания закономерностей наследования признаков при половом размножении очень важно знать эти особенности.
У животных в отличие от растений в онтогенезе очень рано обособляются зачатковые клетки, которые впоследствии дают начало половым железам и половым клеткам. Зачатковые клетки делятся митозом и образуют гонии. Сначала они одинаковы у особей разных полов, затем дифференцируются у самцов в сперматогонии, у самок—в оогонии. Дальнейший процесс их формирования идет по-разному и носит название у самцов—сперматогенеза, у самок— оогенеза. На рисунке 2 изображена схема развития мужских и женских половых клеток у животных.
В процессе сперматогенеза клетки проходят четыре фазы: размножение, рост, созревание, формирование. Сперматогонии делятся митозом с сохранением диплоидного числа хромосом (фаза размножения). Затем деления прекращаются, клетка растет и готовится к мейозу. В это время она называется сперматоцит I (первого порядка). Сперматоцит I вступает в фазу созревания и претерпевает мейоз. В результате первого деления (оно называется у животных первое деление созревания и у большинства бывает редукционным) образуются две гаплоидные клетки, их называют сперматоцитами II (второго порядка). Они делятся еще раз (второе деление созревания, эквационное, уравнительное) и образуют так называемые спер-матиды. Таким образом, из одного сперматогония образуются четыре сперматиды, которые вступают в фазу формирования сперматозоида, т. е. зрелой мужской половой клетки. Сперматозоиды животных имеют иногда очень сложное строение, необходимое для обеспечения им возможности слияния с женской половой клеткой (оплодотворение) в той или иной внешней среде. Сперматозоид состоит из головки, внутри которой помещается ядро, шейки и хвостика.
Сперматогенез у животных, и в частности у млекопитающих, начинается с момента закладки половых желез в эмбриогенезе. Затем после рождения самца он прекращается и вновь возобновляется после полового созревания и протекает постоянно в течение всего периода зрелости. Процесс этот проходит в половых органах—семенниках. Зрелые сперматозоиды по выходе из семенника под влиянием многочисленных гормонов в специальных органах (придатках) приобретают устойчивость к неблагоприятным факторам среды.
В оогенезе (рис. 2) клетка проходит в основном те же фазы, что и в сперматогенезе, но существует целый ряд особенностей. Так, после прекращения делений ооцит I (диплоидная клетка) в отличие от сперматоцита I проходит более выраженную стадию роста. В это время в цитоплазме ооцитов откладывается запас питательных веществ, необходимых для развития зародыша, вследствие чего ооциты приобретают очень большие размеры (например, желток куриного яйца).
После этого ооцит I вступает в мейоз, так же как и сперматоцит I. В результате первого деления созревания образуются две гап-лоидные клетки, но резко отличающиеся друг от друга. Одна, крупная, сохраняет весь запас питательных веществ, называется ооцит II, другая, значительно меньшая, неполноценная, представляет собой выделившееся под оболочку первой клетки ядро. Ее называют редукционное тельце (или полярное, или направительное тельце, или оотида). Она дегенерирует. Иногда перед дегенерацией редукционное тельце успевает разделиться еще раз, образуя два редукционных тельца, но судьба их всегда одинакова - они дегенерируют. Ооцит II делится (второе деление созревания) и образует опять две неравноценные клетки: одна—зрелая яйцеклетка, имеющая гаплоидный набор хромосом и несущая весь запас питательных веществ, другая—второе направительное тельце. Таким образом, в отличие от сперматогенеза из одного оогония образуется только одна функционирующая яйцеклетка, остальные три гибнут. Если вспомнить, что в процессе мейоза образуются клетки, несущие различные комбинации отцовских и материнских хромосом, то это обстоятельство должно иметь существенное значение в определении характера наследования признаков при половом размножении, так как у самки из всех образующихся комбинаций жизнеспособна только одна. Расхождение хромосом в яйцеклетку и направительное тельце в мейозе, как правило, носит случайный характер, поэтому при учете большого числа яйцеклеток сохранение лишь одного из четырех продуктов мейоза не меняет общей картины расщепления.
Яйцеклетка животных обычно круглая, без структурных усложнений, поэтому стадия формирования не играет существенной роли в оогенезе.
Оогенез у млекопитающих происходит в яичниках и тоже начинается в период эмбрионального развития. Так, у женского человеческого эмбриона в возрасте пяти месяцев бывают уже половые клетки на стадии ооцита I. После рождения оогенез вначале приостанавливается (на стадии ооцита I), а затем вновь продолжается с момента полового созревания, яйцеклетки развиваются из ооцитов I, возникших в эмбриогенезе. В течение всего периода половой зрелости оогенез протекает циклически (менструальные циклы у человека, астральные—у животных
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.
Спорогенез, гаметогенез и оплодотворение у растений
У Покрытосеменных растений спорогенез, гаметогенез и оплодотворение представляют собой непрерывную последовательность репродуктивных процессов, происходящих в цветках. В типичных цветках репродуктивные органы представлены гинецеем (совокупностью плодолистиков, образующих женскую репродуктивную сферу) и андроцеем (совокупностью тычинок, образующих мужскую репродуктивную сферу).
Мегаспорогенез, образование зародышевого мешка и яйцеклетки
Мегаспорогенез протекает в женской репродуктивной сфере – в гинецее. Морфологически гинецей представлен пестиком (или несколькими пестиками). В состав отдельного пестика входят: завязь, столбик и рыльце. Внутри завязи находятся семязачатки (один или несколько). Внутреннее содержимое семязачатка называется нуцеллус; снаружи семязачаток окружен покровами (одиночным или двойным интегументом). В нуцеллусе семязачатка (2n) содержится одна археспориальная клетка, способная делиться путем мейоза (у некоторых растений археспорий многоклеточный). В результате мейоза из археспориальной клетки (материнской клетки мегаспор) образуется четыре мегаспоры. Вскоре три из них отмирают, а одна увеличивается в размерах и трижды делится путем митозов. В результате образуется восьмиядерный зародышевый мешок (женский гаметофит). Три ядра вместе с прилегающей цитоплазмой образуют клетки-антиподы, два ядра – одно центральное диплоидное ядро; два ядра – две клетки-синергиды; одно ядро становится ядром яйцеклетки.
Микроспорогенез и образование пыльцевых зерен
Микроспорогенез протекает в мужской репродуктивной сфере – в андроцее. Каждый элемент андроцея – тычинка – состоит из тычиночной нити и пыльника. Внутри пыльника имеются пыльцевые гнезда, содержащие археспориальную ткань. В археспориальной ткани пыльников из каждой диплоидной материнской клетки микроспор в результате мейоза образуется четыре микроспоры. Каждая микроспора делится путем митоза и образуется двухклеточное пыльцевое зерно: одна клетка называется генеративной (в дальнейшем на ее основе образуются спермии), вторая – вегетативной (это остаток вегетативных клеток мужского гаметофита). Зрелое пыльцевое зерно покрыто двойной оболочкой: экзиной (внешней) и интиной (внутренней).
Опыление
Опыление – это процесс переноса пыльцы из пыльников на рыльце пестика. Существует два основных типа опыления: самоопыление и перекрестное опыление. При самоопылении происходит перенос пыльцы в пределах одного цветка, а при перекрестном опылении пыльца из пыльников одного цветка переносится на рыльце пестика другого цветка.
Для более точной характеристики опыления с точки зрения генетики используются специальные термины: клейстогамия (самоопыление в нераскрывшемся цветке), автогамия (самоопыление в раскрывшемся цветке), гейтоногамия (перенос пыльцы из пыльника одного цветка на рыльце пестика другого цветка в пределах одного растения), аллогамия (перенос пыльцы из пыльника цветка одного растения на рыльце пестика цветка другого растения). Таким образом, лишь аллогамия является собственно перекрестным опылением, а остальные типы опыления тождественны самоопылению.
Образование пыльцевой трубки и спермиев
В результате опыления пыльцевое зерно попадает на рыльце пестика и прорастает в пыльцевую трубку (мужской гаметофит). В пыльцевой трубке ядро генеративной клетки делится путем митоза, образуя два генеративных ядра. (У ряда растений деление генеративной клетки происходит еще в пыльниках). Каждое генеративное ядро с прилежащим слоем цитоплазмы называется спермием.
Двойное оплодотворение, образование зародыша и семени
Пыльцевая трубка внедряется в ткани столбика и растет в направлении завязи, при этом на вершине пыльцевой трубки находится вегетативное ядро, а за ним движутся оба спермия. Затем пыльцевая трубка проникает в семязачаток через пыльцевход – микропиле (у некоторых растений существует халазогамия – проникновение пыльцевой трубки в семязачаток через его ножку – халазу). Далее пыльцевая трубка лопается, ее содержимое изливается в зародышевый мешок, и происходит двойное оплодотворение. Один спермий сливается с ядром яйцеклетки, и образуется диплоидная зигота. Другой спермий сливается с центральным диплоидным ядром зародышевого мешка, и образуется триплоидное ядро эндосперма. Все остальные ядра зародышевого мешка и вегетативное ядро пыльцевого зерна – разрушаются.
Из диплоидной зиготы развивается зародыш, а из триплоидного центрального ядра – эндосперм. Происходит формирование семени, включающего зародыш, эндосперм и семенную кожуру, которая образуется из покровов семязачатка.
You can follow any responses to this entry through the RSS 2.0 feed.
You can leave a response, or trackback from your own site.
Читайте также: