
Спорогенез у покрытосеменных растений
Обновлено: 05.10.2024
Особенности гаметогенеза у растений. Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор; 2-й этап — гаметогенез — включает образование зрелых гамет.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) —мега- или макроспорогенезом.
Микроспорогенез протекает аналогично сперматогенезу у животных, вплоть до образования сперматид, а мегаспорогенез соответствует образованию ооцита П.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У растений отсутствует зачатковый путь в том смысле, как это имеет место у животных.
Микроспорогенез и микрогаметогенез. Рассмотрим микроспорогенез и микрогаметогенез на наиболее общем примере покрытосеменных растений. В субэпидермальной ткани молодого пыльника, называемой археспорием, каждая клетка после ряда делений становится материнской клеткой пыльцы, которая проходит все фазы мейоза.
52
В результате двух мейотических делений возникают четыре гаплоидные микроспоры (рис. 24, /). Они лежат четверками, их называют тетрадами спор.
При созревании тетрады распадаются на отдельные микроспоры. Этим заканчивается микроспорогенез.
Вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, не способны к движению и называются спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три клетки. Две из них — спермин и одна вегетативная.
Мегаспорогенез и мегагаметогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, часто только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры (см. рис._ 24, 2).
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады гаплоидна. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития). Судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти, и затем ее ядро претерпевает ряд митозов. При этом сама клетка не делится, она образует зародышевый мешок.
У разных растений число митозов может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) бывает три деления, в результате возникает восемь одинаковых ядер. Во время этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других в противоположном конце зародышевого мешка, называемого халазальным (рис. 25). В дальнейшем ядра обособляются в самостоятельные клетки, имеющие значительное количество цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка и две так называемые синергиды — обра-
53
зуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 одинаковых гаплоидных ядер, из которых только одно образует яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
Кроме моноспорического типа развития, который только что рассмотрен, существуют и другие. При различных типах развития зародышевого мешка сохраняется разное' 1 количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям (если сохраняются две мегаспоры — биспорический; четыре — тетраспорический тип развития).
Сравнение процессов созревания половых клеток у животных и растений (рис. 23 и 24) показывает почти полный параллелизм их, несмотря на то, что расхождение (дивергенция) растений и животных в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов в растительном и животном мире.
ОПЛОДОТВОРЕНИЕ
Сущность оплодотворения. Оплодотворением принято называть побуждение яйца к развитию путем объединения в нем ядер (кариогамии) мужских и женских половых клеток. Оплодотворение представляет собой необратимый процесс; однажды оплодотворенное яйцо не может быть оплодотворено вновь. Син-гамия (слияние мужских и женских половых клеток) и кариогамия составляют сущность процесса оплодотворения.
Оплодотворение у животных. Процесс оплодотворения у животных можно разделить на несколько фаз. Первая фаза начинается с того, что сперматозоид либо прикрепляется к любой точке поверхности яйцеклетки, либо проникает в нее через микропиле. Момент соприкосновения головки сперматозоида с яйцом является начальным в цепи химических реакций. Эту фазу называют фазой активации яйца.
К моменту соприкосновения сперматозоида с яйцом и проникновения его внутрь ядро яйца у разных животных может находиться на разных стадиях деления созревания. Кариогамия же может произойти только после окончания мейоза, т. е. после образования женского пронуклеуса.
В акте оплодотворения два гаплоидных пронуклеуса сливаются в одно ядро, образуя зиготу. Этот момент является кульминационным пунктом процесса полового размножения. В результате кариогамии гомологичные хромосомы, разошедшиеся в мейозе у предыдущего поколения, вновь воссоединяются в одном ядре зиготы. Так восстанавливается диплоидный набор хромосом при половом размножении.
В цитоплазму яйцеклетки у млекопитающих проникает не только головка (ядро) сперматозоида, как это считали раньше, но и его шейка и даже хвостовая часть, что создает возможность передачи некоторого количества цитоплазмы мужского организма потомству.
Оплодотворение у растений. Оплодотворение у растений в принципе сходно с таковым у животных, но имеет и некоторые особенности.
Раньше было сказано, что микрогаметогенез завершается образованием двух спермиев, которые формируются или в пыльцевом зерне, или в пыльцевой трубке при прорастании пыльцевого зерна.
Пыльцевая трубка, дорастая до микропиле зародышевого мешка, соприкасается с яйцевым аппаратом — яйцеклеткой и синергидами (рис. 26, /). При соприкосновении конца пыльцевой трубки с синергидами пыльцевая трубка лопается, а синергиды разрушаются. Передвигающиеся по пыльцевой трубке по мере ее роста два генеративных ядра — спермин после разрыва трубки вместе с ее содержимым попадают внутрь зародышевого мешка (рис. 26, 2). Из двух проникших в зародышевой мешок спермиев один спермин сливается с гаплоидным ядром яйцеклетки (рис. 26, 3). Слияние ядра спермия с ядром яйцеклетки является собственно оплодотворением у растений. В оплодотворенной яйцеклетке — зиготе восстанавливается диплоидное число хромосом. Из зиготы развивается зародыш семени.

ОБРАЗОВАНИЕ ПОЛОВЫХ КЛЕТОК И ОПЛОДОТВОРЕНИЕ У ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ.
Особенности формирования мужских половых клеток
Особенности формирования женских половых клеток
Спорогенез: в основе лежит мейоз (процесс образования гаплоидных клеток); мейозу предшествует митоз (как и у животных)
- протекает в археспориальной ткани
-митотическое деление приводит к образованию
многочисленных материнских клеток пыльцы с
диплоидным набором хромосом, которые вступают в
-после двух мейотических делений образуются
гаплоидные клетки (микроспоры), которые
покрываются двумя защитными оболочками и
превращаются в пыльцевое зерно
- протекает в семязачатках
-обособляется одна, или несколько археспориальных клеток, которые усиленно растут и вступают в мейоз – образуется одна крупная (макроспора) и три мелких клетки (затем разрушаются).
Гаметогенез: предзародышевое развитие — процесс созревания половых клеток, или гамет, в основе лежит митоз.
- протекает в пыльцевом зерне
- в процессе митоза пыльцевое зерно делится и образуется две клетки: вегетативная и генеративная (еще раз делится митозом). В конечном счете образуется два спермия.
-ядро макроспоры делится митозом (три раза), без образования цитокинеза и образуется восьмиядерный зародышевый мешок.
- в дальнейшем формируются самостоятельные клетки: собственно яйцеклетка, две клетки – синергиды, три клетки – антиподы и две, сливаясь, образуют центральную диплоидную клетку.
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру: от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.

Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) - важнейшими производителями органических веществ на суше, как водоросли - в морях и океанах.
Цветок - генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения. Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище - завязи, сформированной из плодолистика (-ов).
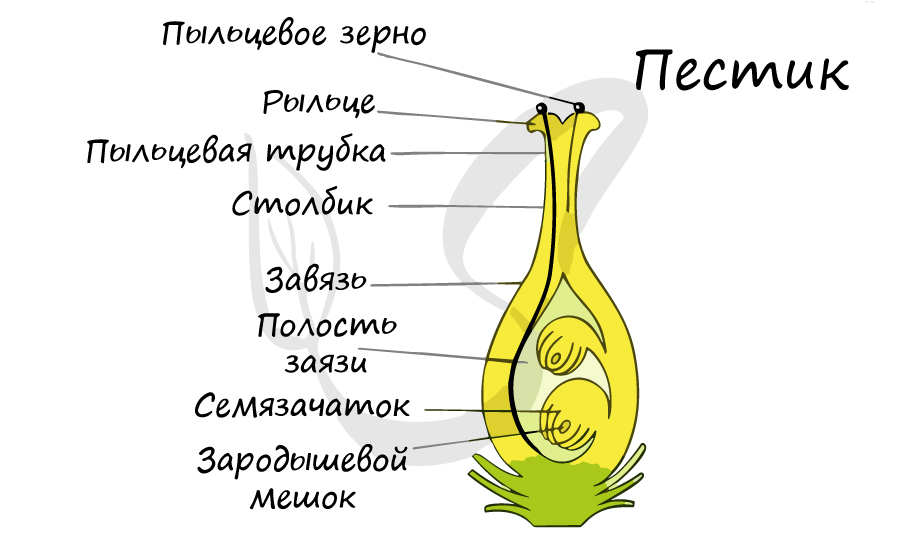
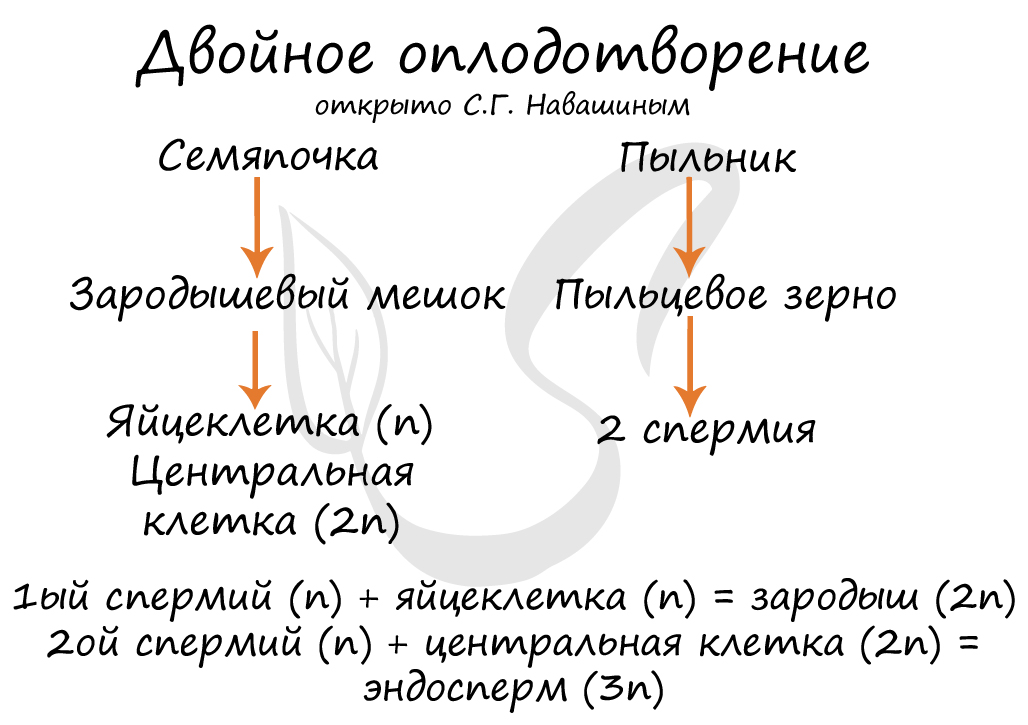
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества - эндосперма (3n). Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод - генеративный орган, служащий для защиты и распространения семян.
Ксилема - проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами, а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

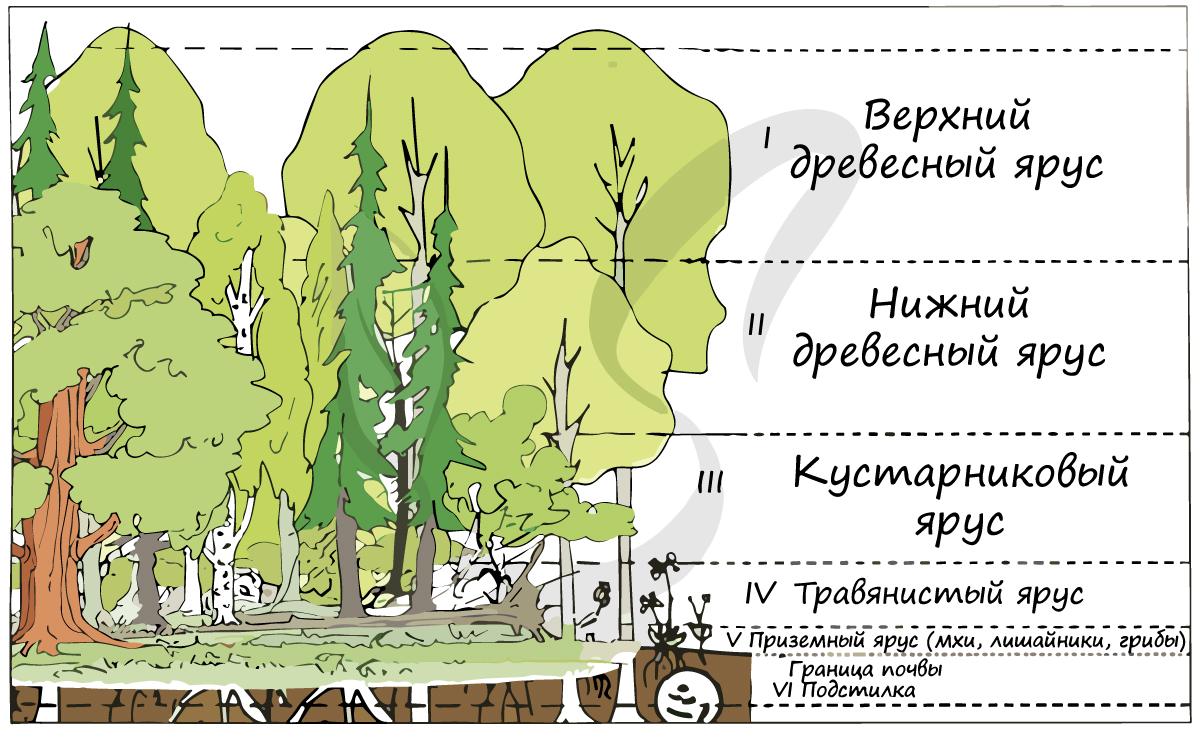
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых :)
Классы покрытосеменных
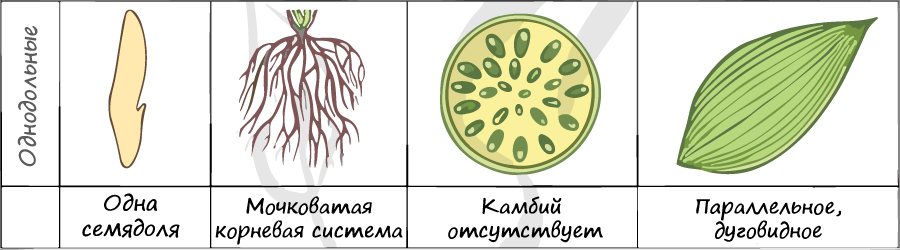
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные, бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
-
Двудольные - семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
-
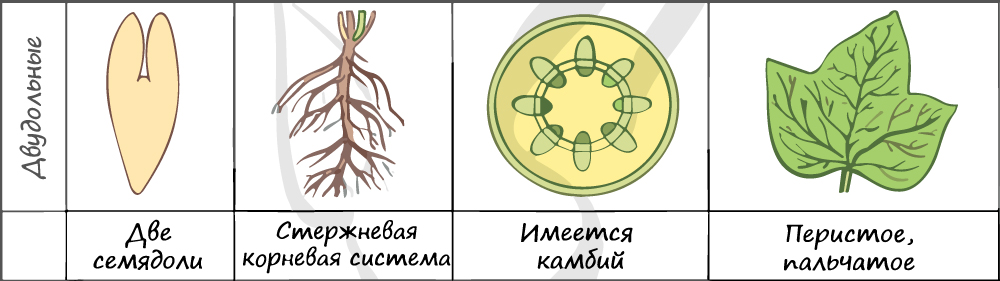
В составе зародыша обычно имеется две семядоли
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни. Главный корень развивается из зародышевого корешка.
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

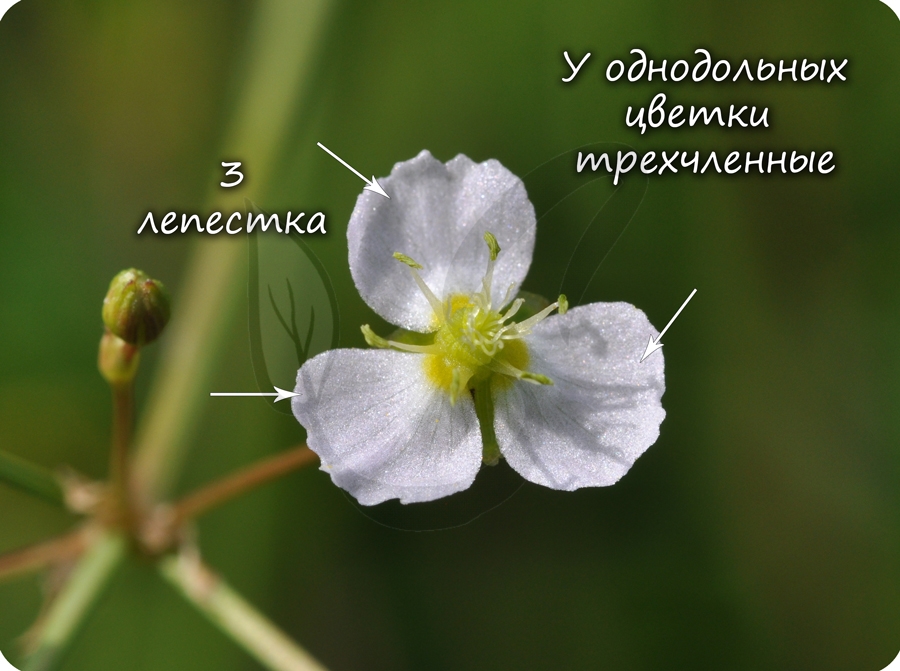
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
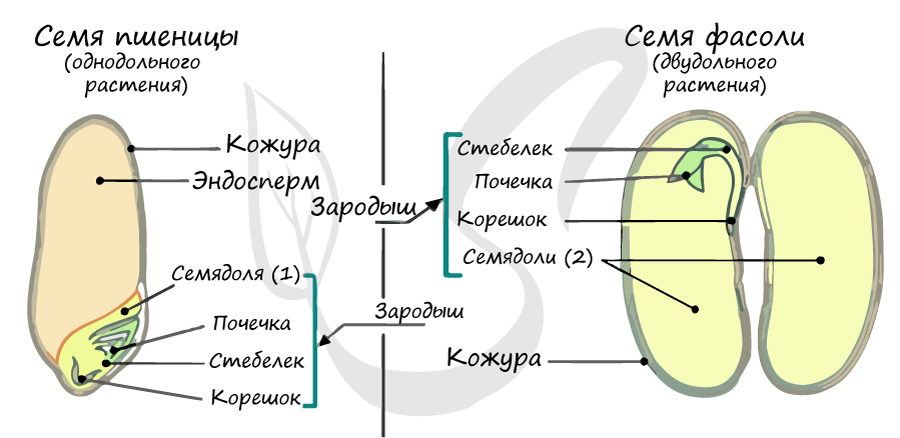
Эндосперм семени
Эндосперм (от греч. endon - внутри + греч. sperma - семя) - запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у - березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
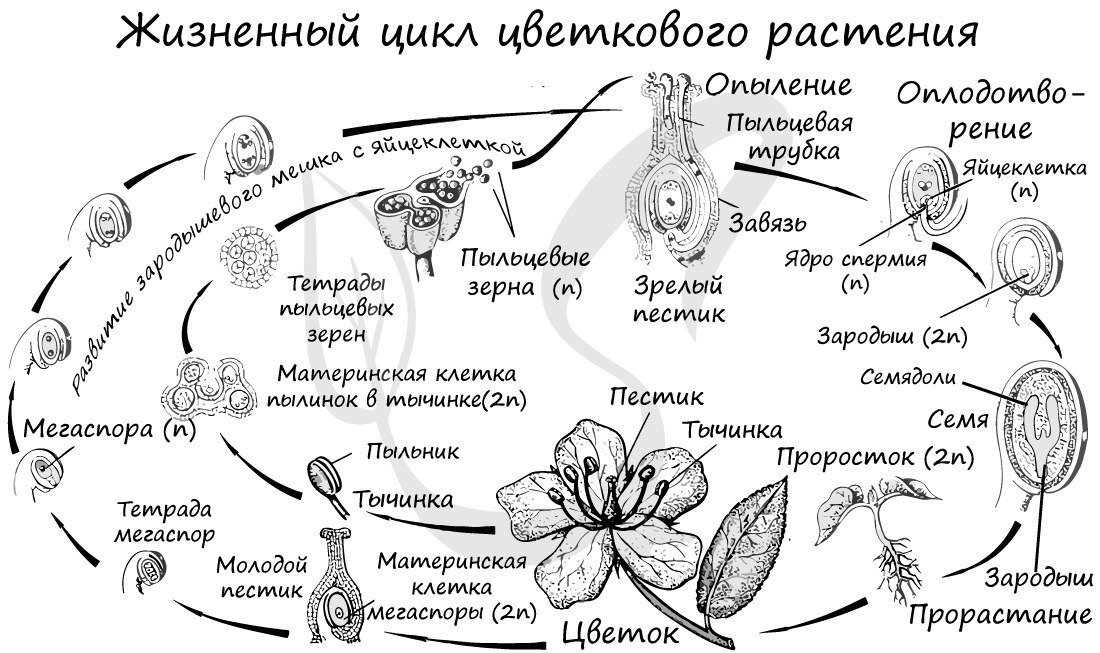
Жизненный цикл
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n) в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) - мужской гаметофит. В завязи пестика в семязачатке формируется женский гаметофит - зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток. Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) - запасного питательного вещества. Другой спермий (n) сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник - образуется плод. Своим внешним видом плоды привлекают животных, и те их охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия, они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте - почти все культурные растения принадлежат к этому отделу! Цветковые имеют медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Микроспорогенез протекает аналогично сперматогенезу у животных, вплоть до образования сперматид, а мегаспорогенез соответствует образованию ооцита И.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У растений отсутствует зачатковый путь в том смысле, как это имеет место у животных.
В субэпидермальной ткани молодого пыльника, называемой археспорием, каждая клетка после ряда делений становится материнской клеткой пыльцы, которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Они лежат четверками, их называют тетрадами спор.
При созревании тетрады распадаются на отдельные микроспоры. Этим заканчивается микроспорогенез.
Вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, не способны к движению и называются спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три клетки. Две из них — спермии и одна вегетативная.
Мегаспорогенез и мегагаметогенез. В субзпидермальном слое молодой семяпочки обособляется археспориальная клетка, часто только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоу.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады гаплоидна. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития). Судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти, и затем ее ядро претерпевает ряд митозов. При этом сама клетка не делится, она образует зародышевый мешок.
У разных растений число митозов может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) бывает три деления, в результате возникает восемь одинаковых ядер. Во время этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других в противоположном конце зародышевого мешка, называемого халазальным. В дальнейшем ядра обособляются в самостоятельные клетки, имеющие значительное количество цитоплазмы. Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка и две так называемые синергиды — образуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 одинаковых гаплоидных ядер, из которых только одно образует яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
Кроме моноспорического типа развития, который только что рассмотрен, существуют и другие. При различных типах развития зародышевого мешка сохраняется разное'1 количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям (если сохраняются две мегаспоры — биспорический; четыре — тетраспорический тип развития).
В завязь покрытосеменных растений проникает два спермия, один из них сливается с яйцеклеткой, дав начало диплоидному зародышу. Другой соединяется с центральной диплоидной клеткой. Образуется триплоидная клетка, из которой возникнет эндосперм - питательный материал для развивающегося зародыша .Этот процесс, характерный для всех покрытосеменных, открыт в конце прошлого века С.Г. Навашиным и получил название двойного оплодотворения. Значение двойного оплодотворения,заключается в том, что обеспечивается активное развитие питательной ткани уже после оплодотворения. Поэтому семяпочка у покрытосеменных не запасает питательных веществ впрок и, следовательно, развивается гораздо быстрее, чем у многих других растений, например у голосеменных.
(1/2 B + 1/2 b) X (1/2 B + 1/2 b) = (1/4 BB + 1/4 Bb + 1/4 bB + 1/4 bb)=3/4 с домин и 1/4рецес.
яйцеклетки сперматозоиды сочетание при оплодотворении
Читайте также: