
Спорогон у мхов это
Обновлено: 07.07.2024
Моховидные, или Мхи, или Настоящие мхи, или Бриофиты (лат. Bryophyta) — отдел высших растений, насчитывающий около 10 000 видов, объединённых примерно в 700 родов и 110—120 семейств (общее число всех мохообразных, включая Печёночные мхи и Антоцеротовые мхи, составляет около 20 000 видов). Как правило, это мелкие растения, длина которых лишь изредка превышает 50 мм; исключение составляют водные мхи, некоторые из которых имеют длину более полуметра, и эпифиты, которые могут быть ещё более длинными. Моховидные, как и другие Мохообразные, отличаются от других высших растений тем, что в их жизненном цикле гаплоидный гаметофит преобладает над диплоидным спорофитом.
Ранее в этот отдел, помимо класса листостебельных мхов, или собственно мхов (в ранге класса — Bryopsida), включали также Печёночные мхи (в ранге класса — Marchantiopsida, или Hepaticopsida) и Антоцеротовые мхи (в ранге класса — Anthocerotopsida), но сейчас эти таксоны повышены до уровня собственных отделов Marchantiophyta и Anthocerotophyta. Для обозначения совокупности этих трёх отделов используется неформальный собирательный термин мохообразные (Bryophytes).
Раздел ботаники, предметом изучения которого являются мохообразные, называется бриологией.
История.
Многие данные указывают на то, что мхи, как и другие споровые растения, произошли от псилофитов (риниофитов) — группы древних вымерших наземных растений. При этом спорофит мхов рассматривается как конечный результат редукции разветвлённого спорофита предковых форм.Есть и другая точка зрения, согласно которой мхи, плауновидные и риниофиты имеют общее происхождение от ещё более древней группы растений.Ранние палеонтологические находки датируются концом девона — началом карбона.
Биологическое описание
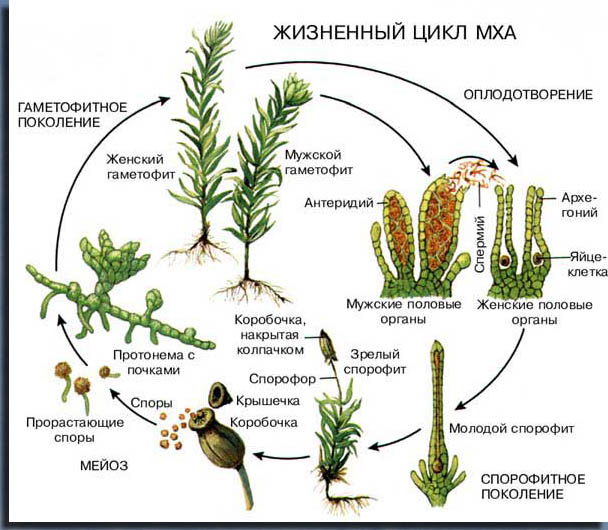
У мхов нет цветков, корней и проводящей системы. Размножаются мхи спорами, которые созревают в спорангиях на спорофите. В жизненном цикле, в отличие от сосудистых растений, преобладает гаплоидный (то есть с одинарным набором непарных хромосом) гаметофит (половое поколение). Гаметофит мхов — многолетнее зелёное растение, нередко с листоподобными боковыми выростами и корнеподобными выростами (ризоидами), в то время как спорофит (или бесполая стадия жизненного цикла) короткоживущий, быстро усыхает и состоит только из ножки и коробочки, в которой созревают споры.
Спорофит моховидных (носящий название спорогоний, или спорогон), имеет более простое строение, чем у других групп высших растений. Он не способен укореняться и располагается на гаметофите. Спорофит состоит, как правило, из трёх элементов:
- коробочка (или спорангия), в которой развиваются споры;
- ножка (или спорофора), на которой располагается коробочка;
- стопы, обеспечивающей физиологическую связь с гаметофитом.
Хлоропласты в клетках Plagiomnium affine из семейства Mniaceae
Со мхами не следует путать так называемый Олений мох (Ягель), поскольку олений мох — собирательное название для нескольких лишайников.
Распространение
Мох, покрывший камни
Мхи встречаются на всех континентах, в том числе в Антарктиде, нередко в экстремальных условиях обитания.
Обычно мхи образуют плотные скопления в затенённых местах, нередко в непосредственной близости от воды, хотя могут встречаться и на относительно открытых, сухих участках. Среди моховидных есть виды, живущие в пресноводных водоёмах. Морских обитателей нет, хотя некоторые виды поселяются на скалах в полосе прибоя.

Значение мхов
- Участвуют в создании особых биоценозов, особенно там, где почти сплошь покрывают почву (тундра).
- Моховой покров способен накапливать и удерживать радиоактивные вещества.
- Играют большую роль в регулировании водного баланса ландшафтов, так как способны впитывать и удерживать большое количество воды.
В деятельности человека:
- Могут ухудшать продуктивность сельскохозяйственных земель, способствуя их заболачиванию.
- Предохраняют почву от эрозии, обеспечивая равномерный перевод поверхностного стока вод в подземный.
- Некоторые сфагновые мхи применяются в медицине (в качестве перевязочных средств при необходимости).
- Сфагновые мхи — источник образования торфа.
Классификация
Листостебельные представители моховидных имеют ряд признаков, которые отличают их от остальных высших растений.

У них отсутствуют корни; прикрепление к субстрату (частично и всасывание воды) осуществляется одним или многочисленными ризоидами — выростами эпидермы. Поглощение воды происходит главным образом нижней частью стебля или телом всего растения (сфагновые мхи).Вегетативные органы сформированы ассимиляционной, проводящей, запасающей и покровной тканями. Однако в отличие от других высших растений у моховидных отсутствуют настоящие сосуды и механические ткани. На стебле зеленых мхов по спирали расположены бесчерешковые листья, имеющие цельную пластинку с жилкой или без нее.Это единственная группа растений, эволюция которой связана с регрессивным развитием спорофита и преобладанием в жизненном цикле гаметофита. Спорофит моховидных —спорогон — состоит из коробочки и ножки, нижняя расширенная часть которой — стопа -— внедряется в ткань гаметофита и доставляет спорофиту воду и питательные вещества. Спорофит, таким образом, лишен самостоятельности и полностью зависит от гаметофита.
Основная функция спорофита — образование спор, которые в благоприятных условиях прорастают. С момента прорастания споры начинается развитие гаметофита — полового поколения моховидных. Прежде всего развивается разветвленная нитчатая (у большинства мхов) или пластинчатая (у сфагнума) протонема, на которой закладываются почки. У одних мхов из почек протонемы формируются пластинчатые слоевища, у других — листостебельные побеги гаметофита (взрослого мха), на которых развиваются органы полового размножения — антеридии и архегонии.
Самую многочисленную группу моховидных составляет класс Настоящие мхи (около 10 тыс. видов), включающий зеленые и сфагновые мхи.
Зеленые мхи.
Они встречаются повсеместно: произрастают на почве, стволах деревьев, скалах и крышах домов, но наиболее широко распространены в сырых лесах, образуя сплошной зеленый ковер. Один из наиболее известных и самых распространенных мхов — кукушкин лен, образующий густые дерновинки из прямостоячих неветвистых стеблей, густо покрытых узкими линейно-ланцетными листьями.Архегонии и антеридии формируются на верхушках стеблей разных особей, которые обычно растут рядом. В антеридиях созревают двухжгутиковые сперматозоиды, в архегониях — по одной неподвижной яйцеклетке. Оплодотворение происходит во время дождя или при обильной росе, поскольку сперматозоиды могут подплыть к архегонию только в воде. Из зиготы здесь же, на верхушке женского гаметофита, развивается спорофит .Внутри коробочки спорогона размещается спорангий, в котором образуются гаплоидные споры. После созревания споры высыпаются из коробочки, разносятся ветром и в благоприятных условиях прорастают, образуя протонему в виде зеленой разветвленной нити. На ней закладываются почки, из которых со временем развиваются листостебельные растения мха.Зеленые мхи способны также к вегетативному размножению молодыми побегами, почками, листьями, выводковыми телами.

Сфагновые мхи.
К сфагновым (белым, или торфяным, мхам) относится примерно 350 видов, составляющих один род сфагнум. Они распространены по всему миру, но наиболее обильно представлены на верховых болотах, где господствуют над всеми остальными видами растений.Строение сфагновых мхов отличается рядом особенностей. У них отсутствуют ризоиды, поэтому вода с растворенными минеральными солями поступает непосредственно в клетки листа и стебля. Стебель гаметофита несет мутовки ветвей, густо покрытых листьями, которые на верхушке главной оси образуют розетку в виде головки почковидной формы. Некоторые боковые, более длинные веточки свешиваются вдоль стебля и плотно прилегают к нему, создавая своеобразную проводящую систему наподобие фитиля, по которому поднимается вода.
Листья сфагновых мхов не имеют средней жилки. Все они состоят из двух типов клеток: живых — ассимилирующих, длинных и узких, с хлоропластами, и мертвых (без протопласта), с утолщениями на стенках и порами. Подобные клетки без живого содержимого присутствуют и в стебле. Благодаря такому анатомическому строению стебля и листа сфагнум способен впитывать и удерживать большое количество воды, которая в 30— 40 раз превышает массу самого растения. Поэтому почва, где поселяются эти мхи, постепенно переувлажняется и заболачивается.
Антеридии и архегонии сфагнумов образуются либо на разных особях, либо на разных веточках одного и того же растения. После оплодотворения из зиготы формируется шаровидная коробочка на ложноножке, являющейся частью гаметофита. При вскрытии коробочки зрелые споры разлетаются на расстояние до 10 см и прорастают, образуя пластинчатую протонему с ризоидом. Из почки на протонеме вырастает гаметофит.
Особенностью сфагновых мхов является беспрерывное нарастание стебля верхушкой и отмирание нижней части. Однако полного сгнивания отмерших частей не происходит, так как в пере-увлажненной почве мало кислорода, а вследствие выделения сфагнумом специфических органических кислот создается кислая среда, в которой прекращается развитие почвенных микроорганизмов (бактерий, грибов и др.), разлагающих растительные остатки. В течение длительного времени происходит накопление значительных запасов органических веществ в виде торфа. Процесс образования торфа идет очень медленно: слой торфа в 1 см накапливается за 10 лет, толщиной в 1 м — за тысячу лет.
СПОРОГОН — спорофит (бесполое поколение) мохообразных, состоящий из коробочки, ножки и присоски (гаустория), внедряющийся в ткань гаметофита (половое поколение) в получающий от него питательные вещества. СПОРОГОРМОГОНИИ, ПЕРВИЧНЫЕ ГОРМОГОНИИ — проростки спор сине-зеленых водорослей из кл. Hormogoniophyceae, имеющие гормогониеподобное строение.
Словарь ботанических терминов. — Киев: Наукова Думка . Под общей редакцией д.б.н. И.А. Дудки . 1984 .
Смотреть что такое "СПОРОГОН" в других словарях:
спорогон — а, ч. Те саме, що спорогоній … Український тлумачний словник
спорогон — см. спорогоний … Анатомия и морфология растений
спорогоній — я, ч. Нестатеве покоління (спорофіт) мохоподібних … Український тлумачний словник
Спорогоний — спорогон (от Споры и греч. gónos рождение, плод), Спорофит (бесполое поколение) мохообразных, состоящий из коробочки, ножки и присоски (гаустория), внедряющейся в ткань Гаметофита (половое поколение) и получающей от него питательные… … Большая советская энциклопедия
Мхи — наземные или реже пресноводные автотрофные растения, объединяемые в отдел мохообразных (См. Мохообразные), включающий наиболее примитивные высшие растения. М. подразделяют на 3 класса: антоцеротовые, печёночные и лиственные, или… … Большая советская энциклопедия
Развитие и строение спорофита — Спорофит образуется из оплодотворенной яйцеклетки в ткани слоевища. По сравнению с низкоорганизованным, слабо расчлененным гаметофитом спорофит у антоцеротовых относительно высоко развит. У большинства представителей этого класса… … Биологическая энциклопедия
Спорофит — Как и у всех моховидных, диплоидный спорофит (спорогон) печеночников прикреплен к гаметофиту и осуществляет свое питание за счет фотосинтетической деятельности его тканей. Клетки спорогопа у большинства печеночников не содержат… … Биологическая энциклопедия
Лиственные мхи — листостебельные мхи (Bryatae, Musci), класс споровых растений отдела мохообразных. Тело Л. м., в отличие от др. мхов, расчленено на стебель и листья. Класс объединяет 3 подкласса: сфагновые (Sphagnidae), андреевые (Andreaeidae) и бриевые… … Большая советская энциклопедия
Общая характеристика1 — Из всех моховидных листостебельные мхи включают наибольшее число видов. Среди них многие отличаются широким распространением (иногда в умеренных и холодных зонах обоих полушарий), высокой жизненной устойчивостью и большой фитоценотической … Биологическая энциклопедия
мхи — (моховидные, бриофиты), отдел высших растений. Преимущественно многолетние растения, характеризующиеся групповыми формами роста (дерновинки, куртинки, подушки). Органы полового размножения архегонии и антеридии, бесполого спорогон (диплоидный) … Энциклопедический словарь

Целью изучения данной темы является формирование системы знаний о высших споровых растениях как наиболее примитивной группе высших растений, чьё размножение тесно связано с водой, об их роли в эволюции высших растений и значении в природе. Учащиеся сформируют представление о строении тела листостебельных, печёночных и антоцеротовых мхов, рассмотрят жизненный цикл мохообразных и как происходит чередование поколений.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Высшие растения. Отдел Мохообразные"
Мохообразные – единственный отдел высших растений, у которых гаметофит является преобладающим поколением.
Растение, которое мы представляем себе как мох, и является гаметофитом.
Мохообразные отличаются относительно простым строением. Их тело расчленено на стебель и лист, а у наиболее простых тело представляет собой слоевище (таллом).
Однако все вегетативные органы мохообразных не могут считаться истинными, поскольку они принадлежат гаметофиту и негомологичны настоящим органам спорофита.
Мохообразные не имеют корней и прикрепляются к субстрату нитевидными ризоидами, которые у различных форм могут быть одно или многоклеточными.
Поглощение воды и других необходимых веществ растением осуществляется не только ризоидами, но и через всю поверхность тела. Поглощение осуществляется благодаря полному (или почти полному) отсутствию на поверхности тела защитной кутикулы.
Это вынуждает мхи обживать места с повышенной влажностью. У большинства мохообразных отсутствует проводящая ткань. Кроме того, у них отсутствуют или слабо развиты и большинство других тканей, в частности механические, что ограничивает размеры растений несколькими сантиметрами.
Отдел Мохообразные разделяют на три класса: листостебельные мхи, печёночные, антоцеротовые.
Самый крупный класс – это листостебельные мхи.

Название этого класса отражает его внешний вид и строение – растения состоят из стеблей, которые не ветвятся. Это стебли с покрывающими их выростами-листьями разнообразной формы, расположенными по спирали.
Один из самых известных листостебельных мхов – кукушкин лён – это многолетнее растение. Его часто можно встретить в заболоченных местах. Стройные коричневые стебельки этого мха покрыты маленькими тёмно-зелёными листиками и похожи на миниатюрные растения льна.
В виде подушкообразных дерновин он распространён в лесах, тундре, горной местности, на болотах и лугах, зачастую формирует густой полог.
У кукушкиного льна есть мужские и женские растения. На женских растениях развиваются коробочки на длинных ножках.
В верхней части коробочка прикрыта колпачком с тонкими волосками. Коробочки напоминают сидящую кукушку. Отсюда и название мха – кукушкин лён.
В этих коробочках развиваются споры. Споры высыпаются, а при наступлении благоприятных условий прорастают. Из проросшей споры образуется тонкая зелёная нить – предросток, или протонема. Нить ветвится, на ней появляются почки, из которых затем вырастают мужские или женские экземпляры мха, так осуществляется бесполое размножение.
Половое же размножение кукушкиного льна происходит с участием половых клеток, которые развиваются в половых органах.
В результате слияния половых клеток, образуется клетка (зигота), которая содержит двойной набор хромосом, то есть генетическую информацию от отца и матери. Из этой клетки на женском растении развивается коробочка на ножке – бесполое поколение, из которой будут рассеиваться и прорастать споры.
Среди распространённых видов листостебельных мхов, выделяют и тортулу настенную. Этот мох образует серо-зелёные подушки высотой не более 1 см с язычковидными листьями, имеющими острые или закругленные вершины, которые приближаются к острию. Растёт тортула на известняковых камнях и на других поверхностях из такого материала.
Также к листостебельным мхам относят и широко известный сфагнум – сфагновый (торфяной) мох.
У сфагнума, в отличие от кукушкиного льна, сильно ветвящийся стебель. Также он не имеет ризоидов. Ежегодно мох нарастает верхней частью, а снизу отмирает.
Стебель и ветви большинства видов сфагнума покрыты мелкими светло-зелёными листьями. Имеются ножка и коробочка со спорами.
Для сфагнума характерны особые водозапасающие клетки на листьях и стебле (прозрачные, мёртвые, полые с отверстиями).
Водозапасающие клетки окружены более мелкими зелёными фотосинтезирующими клетками, которые объединены в одну сеть.
В зелёных клетках за счёт фотосинтеза образуются органические вещества, которые поступают из листьев в стебель.
Между зелёными клетками находятся другие, более крупные прозрачные мёртвые клетки. Их цитоплазма разрушена, сохранились только оболочки с отверстиями, поэтому эти клетки прозрачны и могут заполняться водой или воздухом.

Мёртвые клетки листьев и стеблей сфагнума способны поглощать воду в 20–25 раз больше своей массы, долго её удерживать, постепенно отдавая живым клеткам.
Тело сфагнума содержит фенол, который является антисептиком, убивающим бактерии. В связи с этим мох почти не гниёт и образует торф (по 1–2 мм в год). За счёт роста сфагнума и других водных растений происходит заболачивание лесов и зарастание водоёмов: озёра превращаются в болота.
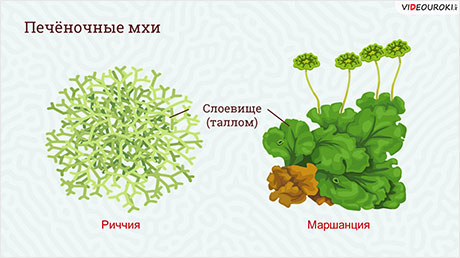
Старейший класс мхов – это печёночные мхи. Большинство мхов-печёночников имеют и стебли, и настоящие листья.
Самый известный представитель этой группы ─ маршанция изменчивая. Это многолетнее растение, имеющее вид зелёного лопастного листка (слоевища), который стелется по поверхности земли.
На верхней поверхности листьев видны чашечки, где находятся выводковые почки (корзиночки), представляющие собой специализированные органы бесполого размножения. Внутри таких корзиночек развиваются многоклеточные выводковые тельца, которые опадают и при прорастании дают начало новому растению.
Ещё один представитель печёночных мхов – риччия. Это плавающее растение, которое зелёным ковром покрывает поверхность воды.
При достаточном количестве света риччия быстро разрастается, образуя плотные подушки на поверхности воды.
Если поместить в банку с влажной почвой немного риччии, а затем накрыть банку стеклом и поставить в тёплое место, то можно наблюдать, как мох быстро разрастается.
Представители класса антоцеротовых мхов имеют тела в виде пластинчатого слоевища.
Представители класса – талломные растения, листостебельные виды отсутствуют. Талломы пластинчатые (лопастные), обычно в форме розеток. По краям талломов возникают клетки образовательной ткани (меристематические клетки), которые формируют лопасти, налегающие друг на друга.
Развитие мохообразных представляет собой чередование гаплоидной и диплоидной фаз. Преобладающим поколением является гаметофит. Он развивается из споры, ядро которой имеет гаплоидный набор хромосом, поэтому все клетки гаметофита тоже гаплоидные. Первоначально из споры развивается протонема, имеющая вид ветвящейся нитчатой водоросли или пластинки.
Спора не содержит запаса питательных веществ, поэтому молодой гаметофит должен их синтезировать самостоятельно посредством фотосинтеза. Наличие необходимой для этого хлоренхимы определяет зелёную окраску гаметофита. У печёночных мхов многократное деление верхушечной клетки даёт начало пластинчатым структурам, на которых впоследствии разовьются половые органы.
У листостебельных мхов гаметофит имеет вид облиственных побегов, они развиваются из почек, которые формируются на протонеме. На гаметофорах образуются половые органы – женские архегонии и мужские антеридии. Чаще всего на одном растении развиваются органы только одного пола – однодомные мхи, но нередко встречаются и двудомные (на одной особи формируются как женские половые органы, так и мужские). Антеридии представляют собой овальные тельца, окружённые тонкой однослойной оболочкой.
Они наполнены сперматогенными клетками, которые при митотическом делении дают начало двум подвижным сперматозоидам, снабжённым двумя жгутиками. Архегоний устроен сложнее и обычно представляет собой структуру, по своей форме напоминающую бутылку.
В утолщённой части, называемой брюшком, находится крупная яйцеклетка, которая также образуется в результате митоза. Внутри суженной шейки в один ряд располагаются шейковые клетки, одна из которых – брюшная шейковая клетка – находится над яйцеклеткой.
Оплодотворение возможно только в капельно-жидкой водной среде. Через шейку сперматозоиды проникают в брюшко архегония и оплодотворяют находящуюся там яйцеклетку. В результате образуется диплоидная зигота, которая по прошествии определённого периода покоя даёт начало диплоидному поколению – спорофиту.
Внутри коробочки многочисленные материнские клетки спор делятся мейотически и дают начало гаплоидным спорам, посредством которых осуществляется бесполое размножение. В коробочке печеночных мхов среди спор находятся пружинки – элатеры. При повышении влажности элатеры раскручиваются, а при понижении вновь скручиваются, при этом споры перемешиваются и разрыхляются, что способствует выталкиванию их из коробочки после того, как она раскрывается. Споры, выпадающие из коробочки, подхватываются ветром и переносятся на большие расстояния.
Подавляющее большинство погибают, попадая в неблагоприятную среду, но количество их столь велико, что часть обязательно попадает во влажную почву, где из них прорастает протонема, дающая начало молодому гаметофиту.
У мохообразных также широко распространено вегетативное размножение. У печёночников образуются выводковые тельца и выводковые корзиночки, а у листостебельных мхов из структур гаметофита могут образовываться вторичные протонемы.
Значение мохообразных в природе в основном связано с участием в почвообразовании. Сами они не испытывают потребности в питательном субстрате, поскольку не обладают корнями и необходимые им вещества способны всасывать всей поверхностью тела.
Поэтому мохообразные одними из первых (наряду с лишайниками) заселяют места, доселе лишённые растительности, постепенно покрывая их сплошным ковром.
Мохообразные очень редко поражаются микроорганизмами, крайне неохотно поедаются насекомыми, птицами и травоядными животными, и, несмотря на медленный рост (несколько миллиметров в год), в итоге они привели к образованию огромных залежей торфа, который образуется из-за неполного гниения постепенно отмирающих нижних частей растений. Чаще всего торф используют в сельском хозяйстве в качестве органического удобрения. Кроме того, он является хорошим топливом и широко применяется в строительной индустрии благодаря высоким теплоизолирующим свойствам.
Мохообразные также участвуют в регуляции водного баланса, так как они задерживают воду и переводят её в состояние грунтовой. Нередко это приводит к заболачиванию почв, что ухудшает их продуктивность.
Бесплатные экскурсии в музей Пиявки!
Международный Центр Медицинской Пиявки приглашает посетить музей и узнать о пользе и вреде пиявок, их выращивании, гирудотерапии, лечебной косметике и многом другом. Подробнее >>>
Зимние учеты птиц России!
Приглашаем биологические кружки, профессиональных орнитологов и просто любителей птиц принять участие в программах зимних учетов птиц "Parus" и "Евроазиатские Рождественские учеты" в зимний сезон 2020-2021 годов. Подробнее >>>
Биологический кружок ВООП приглашает!
Биологический кружок при Государственном Дарвиновском музее г.Москвы (м.Академическая) приглашает школьников 5-10 классов на занятия в музее, экскурсии по вечерам, учебные выезды в природу по выходным и дальние полевые экспедиции в каникулы! Подробнее >>>
Соревнования по полевой ботанике "ВЕСЕННЯЯ ФЛОРА" пройдут в мае-июне 2020 года в онлайн-формате (определение растений по фотографиям). К участию в соревновании приглашаются школьники и взрослые любители природы, проживающие в средней полосе Европейской части России. Подробнее >>>
Здесь может быть бесплатно размещено Ваше объявление о проводимом Всероссийском конкурсе, Слёте, Олимпиаде, любом другом важном мероприятии, связанном с экологическим образованием детей или охраной и изучением природы. Подробнее >>>
Мы публикуем на нашем сайте авторские образовательные программы, статьи по экологическому образованию детей в природе, детские исследовательские работы (проекты), основанные на полевом изучении природы. Подробнее >>>
МХИ: ИЛЛЮСТРИРОВАННЫЙ КОНСПЕКТ ДЛЯ УЧИТЕЛЯ
Отдел Моховидные (Bryophyta).
I. Класс Листостебельные мхи, или Мхи, или Бриопсиды (Bryopsida, или Musci).
Подклассы
1. Sphagnidae. 2. Bryopsidae. 3. Andreidae.
II. Класс Печеночники, или Печеночные мхи (Marchantiopsida, или Hepaticopsida).
Подклассы
1. Jungermanniidae. 2. Marchantiidae.
III. Класс Антоцеротовые, или Антоцеротопсиды (Anthocerotopsida).
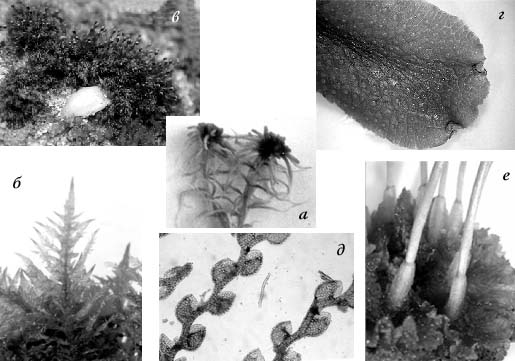
Рис. 1. Представители основных групп мохообразных: а – Sphagnidae; б – Bryopsidae; в – Andreidae; г – Jungermanniidae; д – Marchantiidae; е – Anthocerotopsida.

Рис. 2. Формы роста мхов: а – наземные, б – эпифитные, в – водные, г – полуводные
Мхи – очень маленькие растения и обычно растут там, где влажно. В сырых лесах и безлесных местообитаниях они образуют сплошные ковры. Но есть и виды, способные расти в крайне сухих местообитаниях.
Так же, как и другие растения, мхи:
– являются многоклеточными организмами, способными размножаться половым путем;
– имеют специализированный орган, внутри которого развивается зигота;
– имеют споры и спороносные органы, покрытые кутикулой.
В отличие от других растений, мхи:
– не имеют лигнина в клеточных стенках;
– сильно зависят от влажности воздуха (временно прекращают жизнедеятельность в засушливый период);
– не имеют настоящих корней.
ХАРАКТЕРИСТИКА ОСНОВНЫХ ГРУПП БРИОФИТОВ
Класс Бриопсиды (Bryopsida)
Подкласс сфагновые мхи (Sphagniidae)
Размеры стеблей мхов измеряются в сантиметрах, их листьев – в миллиметрах, а клеток – в микрометрах.


Стебель у сфагнов прямостоячий, но отдельные растения не способны расти прямо из-за отсутствия механической ткани. Поэтому сфагнум всегда растет дернинками (рис. 3), в которых отдельные растения опираются друг на друга веточками, растущими из стебля горизонтально, подобно тому, как могут опираться, обнявшись, друг другу на плечи люди. На верхушке стебля находится более или менее компактная головка, состоящая из скученных вокруг точки роста коротких молодых веточек (рис. 4). По мере роста стебля вверх веточки вырастают в длину и рассредоточиваются по стеблю отдельными пучками, состоящими из горизонтально отстоящих и свисающих вдоль стебля веточек (рис. 5). Отстоящими веточками соседние растения в дернинке сцепляются между собой. По пористым клеткам листьев этих веточек осуществляется горизонтальный транспорт веществ в сфагновой дернинке. По клеткам листьев свисающих ветвей осуществляется транспорт веществ по вертикали (из почвенного раствора). Листостебельное растение сфагна не имеет ризоидов.
Рис. 5. Пучок веточек у сфагна
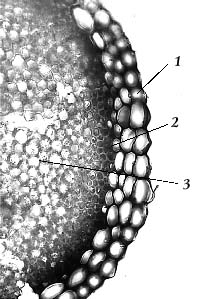
Гаметофор сфагнов состоит из двух типов клеток – живых, хлорофиллоносных, и мертвых, лишенных содержимого крупных водоносных, или гиалиновых, клеток (рис. 6). Стенки последних укреплены спиральными волокнами, предохраняющими их от разрыва при потере тургора. У многих видов в стенках гиалиновых клеток имеются поры, облегчающие всасывание воды (рис. 6, 3). Так как листья, произрастающие на веточке, налегают друг на друга, как черепица на крыше дома (их так и называют – черепитчато-налегающими), поры клеток соседних листьев накладываются друг на друга, так что в олиственной веточке образуется капиллярная система. Она продолжается между веточками и соседними растениями. Толщина листьев сфагновых мхов – всего лишь в один слой клеток (рис. 7).
Более сложное строение обнаруживает стебель (рис. 8). В нем клетки, хотя и дифференцированны, но не образуют специализированных тканей, как у сосудистых растений. Внутренняя часть стебля заполнена тонкостенными клетками, осуществляющими как транспорт веществ к точке роста, так и запасающую функцию. Совокупность этих клеток называется сердцевиной стебля. Сердцевина окружена узкими толстостенными клетками склеродермиса, выполняющего функцию коры и частично – механическую функцию. Кнаружи от склеродермиса имеется слой мертвых тонкостенных гиалиновых клеток, образующих гиалодермис. В гиалодермисе накапливаются и сохраняются запасы влаги, осуществляется траспорт воды к точке роста.
Спорогон сфагнов, как и у других мхов, состоит из ножки и коробочки (рис. 9), но в отличие от класса Bryopsida, ножка спорогона очень короткая (ее функцию выполняет ложноножка, образующаяся из гаплоидных клеток брюшка архегония), а коробочка не имеет перистома.

Рис. 10. Стеблевой (1) и веточные (2) листья сфагна
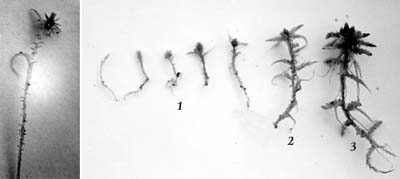
Рис. 11. Онтогенез гаметофита у сфагнов: 1 – изофильные растения; 2 – гемиизофильное растение; 3 – анизофильное растение
Экологические особенности и распространение
Мхи способны заселять наиболее бедные и влажные местообитания. В умеренной зоне и в Арктике они образуют обширные торфяные болота. Торф – это неразложившиеся спрессованные остатки сфагновых мхов, которые сохраняются практически в неизменном виде столетиями и тысячелетиями. Многие болота таежной зоны имеют возраст от 8 000 до 10 000 лет. Вместе со сфагнами гнению не подвергаются и остатки других болотных растений. Происходит это потому, что торфяные мхи выделяют в окружающую среду целый букет органических кислот (яблочную, лимонную, уксусную, муравьиную и др.). В такой кислой среде из организмов, питающихся растительным опадом, способны существовать только некоторые виды грибов. Но и их деятельность подавляется очень сильным фунгицидом – сфагнолом (гликозидом с фенольной основой, химическая формула которого до сих пор точно не известна), который тоже выделяют в почвенный раствор сфагновые мхи. Он же оказывает и гербицидное действие на всходы древесных растений. Поэтому на старых торфяных болотах часто нет леса.
Но сфагны способны расти не только в местообитаниях с бедным минеральным питанием. Часто их можно встретить и в поймах лесных речек, и на озерах, и у родников (выходах ключевых вод). В таких местообитаниях они, правда, не образуют сплошных ковров, а также торфа, так как биохимический состав растений тех видов мхов, которые здесь произрастают, не столь химически агрессивен.
В подобных местообитаниях сфагны произрастают преимущественно в тропической и субтропической зонах, за исключением торфяных болот Бразилии и Новой Зеландии. Собственно, субтропики являются родиной сфагновых мхов, что видно как по наличию в этой зоне примитивных форм, так и по разнообразию видов. Так, во всем мире насчитывается около 300 видов рода Sphagnum, из которых в умеренной зоне Евразии и Америки произрастает только около 50 (рис. 12). Несмотря на более упрощенное строение, по сравнению с представителями класса Bryopsida, сфагны эволюционно более молодые.
Рис. 12. Ареал сфагновых мхов
Подкласс Настоящие мхи (Bryopsidae )
В нижней части стебля у бриевых мхов имеются нити из лишенных хлорофилла клеток – ризоиды, которые выполняют функцию закрепления на субстрате и проведения воды к стеблю (рис. 13). Ризоиды образуются на ранней стадии прорастания споры, параллельно с протонемой. Протонема может быть нитчатой (похожей на водоросли) и пластинчатой (рис. 14). Протонема с почками возобновления листостебельных растений (поздняя стадия развития протонемы) называется каулонемой (рис. 15).
Рис. 15. Каулонема (протонема с почками)
Стебель имеет относительно сложное строение (рис. 16, 17). Внешний слой образует эпидермис, из клеток которого могут вырастать многоклеточные нити – ризоиды. Следующий за эпидермисом слой называется склеродермис. Он состоит из узких и длинных толстостенных клеток, выполняющих проводящую и механическую функции. Самый внутренний слой клеток называется центральным пучком, он состоит из сосудов, очень напоминающих сосуды других высших растений. Различают два типа клеток сосудов – лептоиды (похожие на клетки флоэмы) и гидроиды (подобные клеткам ксилемы). Совокупность лептоидов в стебле называют лептом, а гидроидов – гидром (рис. 18). Помимо лептоидов и гидроидов в центральном пучке имеются стереиды, сходные с волокнами склеренхимы сосудистых растений. В месте прикрепления стеблевого листа к стеблю от центрального цилиндра ответвляется пучок лептоидов и гидроидов, образующих листовую вставку (аналогичную листовому следу у сосудистых). Анатомия стебля сильно варьирует у разных представителей бриофитов, у конкретных видов в состав стебля могут входить не все перечисленные компоненты или они могут по-разному комбинироваться.
Рис. 20. Лист Bryopsidae с центральной жилкой
Жизненный цикл
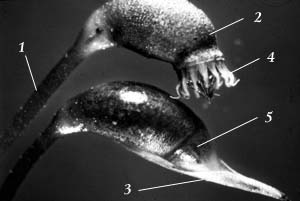
Рис. 24. Спорогон: 1 – ножка; 2 – коробочка; 3 – колпачок; 4 – перистом у открытой коробочки; 5 – крышечка
Спорофит вырастает примерно через месяц–другой после оплодотворения. Внутри незрелой коробочки содержится спорогенная ткань, состоящая из гаплоидных клеток. Зрелым спорогон становится после мейотического деления клеток спорогенной ткани, в результате которого образуются гаплоидные споры.
Вегетативное размножение
Половое размножение у мхов изучено достаточно хорошо. Однако существует множество видов, которые вообще никогда не производят спорогонов. Вегетативный способ размножения характерен для всех видов мхов без исключения. Новое листостебельное растение может развиться буквально из любого фрагмента гаметофита, даже из единственной клетки, а также (у некоторых видов) из клеток ножки спорофита.
Кроме этого имеются и специализированные органы для вегетативного размножения. Это выводковые почки и тельца (геммы) (рис. 25).
Рис. 25. Выводковые тельца мхов (геммы): внизу – прорастающее выводковое тельце с ризоидом
В зависимости от того, в какой части гаметофита образуется спорофит, листостебельные мхи делят на верхоплодные и бокоплодные. Верхоплодные мхи имеют прямостоячий гаметофор, как правило, без боковых веточек. Спорофит у них вырастает из верхушки стебля (по крайней мере, верхней его части). Бокоплодные мхи имеют многочисленные боковые веточки, главный стебель не прямостоячий, а полегающий. Спорогоны образуются в латеральных частях стебля или на верхушках боковых веточек (рис. 26).
Рис. 26. Верхоплодные (1) и бокоплодные (2) мхи
Подкласс андреевые мхи (Andreidae )
Андреевые мхи – это группа наскальных мхов (рис. 1, в). Они способны поселяться на монолитной каменистой породе, буквально вгрызаясь в каждую трещинку ризоидами. Формируют довольно плотные дерновинки. В клеточных стенках листьев андреевых мхов находится красный пигмент, благодаря чему растения имеют темно-красный цвет. Этот пигмент выполняет функцию светофильтра для хлоропластов, которые у этих мхов не способны нормально фунционировать при полном освещении.
В мире насчитывается около 100 видов андреевых мхов, большинство из которых произрастают в полярных регионах северного и южного полушарий. Некоторые виды растут в альпийском поясе гор тропиков.
Рис. 27. Растение андреевого мха с раскрытой коробочкой
Andreidae по некоторым признакам сходны со сфагновыми мхами, по некоторым – с печеночными и бриевыми. Так, их листостебельный гаметофит по внешнему виду напоминает гаметофит кукушкина льна (род Polytrichum). Так же, как и у бриевых мхов, листья имеют центральную жилку. Так же, как сфагновые мхи, андреевые имеют пластинчатую протонему. Коробочка андреевых мхов раскрывается на несколько створок (рис. 27), как у печеночников, но в отличие от последних, верхние концы створок остаются соединенными друг с другом, что придает созревшему спорогону вид японского фонарика. Так же, как и у сфагновых мхов, у андреевых ножка спорогона очень укорочена, так что коробочка выносится вверх с помощью ложноножки, образующейся из гаплоидных тканей архегония.
Класс печеночные мхи (Hepaticae )
В мире насчитывается около 8500 видов печеночников, которые распространены во всех климатических зонах – от тропиков до полярных широт. Хотя среди них имеются виды, способные расти в крайне засушливых местообитаниях или в водоемах, большинство все же встречается в умеренно увлажненных, таких как сырые луга, берега речек.
Преобладающим поколением в жизненном цикле печеночников является гаметофит, который может быть как олиственным (подкласс Юнгерманниевые), так и талломным (подкласс Маршанциевые). Размеры его у разных видов колеблются от 0,15 до 25 см (рис. 28, 29).


Вегетативное размножение печеночников осуществляется путем ветвления стебля (вновь образовавшаяся веточка отделяется от материнского растения и продолжает рост самостоятельно) или с помощью специализированных выводковых органов. Последние представляют собой корзиночки с выводковыми почками (рис. 31). По внешнему виду эти корзиночки могут быть серповидными, чашевидными или бутылковидными
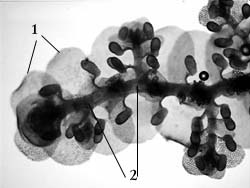
.
Рис. 30. Юнгерманниевый печеночник: 1 – листья; 2 – амфигастрии

Генеративное размножение осуществляется специализированными половыми органами – антеридиями и архегониями (рис. 32). Оплодотворение осуществляется в жидкой среде. Из оплодотворенной яйцеклетки вырастает спорофит, состоящий из коробочки со спорами на ножке (рис. 33, 1)

Рис. 33. Спорофит печеночников: 1 – незрелый; 2 и 3 – зрелый, после рассеивания спор
Зрелая коробочка раскрывается на четыре створки, благодаря чему происходит рассеивание спор (рис. 33, 2, 3). Спора дает начало новому гаметофиту.
Класс антоцеротовые (Anthocerotopsida)
Антоцеротовые – самый маленький класс в отделе Bryophyta. Виды этого класса распространены по всему земному шару и произрастают во влажных тенистых местообитаниях. Гаметофит внешне похож на таллом маршанциевых. Спорофит имеет апикальную меристему и поэтому постоянно растет вверх. Говорят, что он напоминает рога (рис. 34). Споры созревают, начиная с нижней его части. Зрелый спорофит раскрывается сразу по всей длине.
Читайте также: