
Тропические листопадные леса редколесья и кустарники описание
Обновлено: 07.07.2024
Листопадные леса умеренной зоны (широколиственные леса) расположены южнее тайги, не имеют сплошного распространения. Климат здесь более мягкий, осадков 700— 1500 мм/г, с четко выраженными сезонами. Среди древесной флоры доминируют бук и дуб. Структура этих лесов многоярусная: до трех ярусов деревьев, двух ярусов кустарников и двух-трех ярусов трав. Опад листьев формирует мощную лесную подстилку, позволяющую перезимовать многим животным. Богата и разнообразна орнитофауна. Крупные животные здесь те же, что и в тайге. Среди насекомых много вредителей лесов.[ . ]
Листопадные леса умеренной зоны (широколиственные леса) (Западная Европа, Восточная Азия, восток США). Климат сезонный с зимними температурами ниже 0° С, осадков 750— 1500 мм в год. Растительность: господствуют широколиственные листопадные породы деревьев (дуб, бук, клен, липа, ясень, граб), кустарниковый подлесок, мощная лесная подстилка. Животный мир , млекопитающие (лоси, медведи, рыси, лисицы, волки, белки, землеройки), птицы (дятлы, дрозды, совы, соколы). Биота адаптирована к сезонному климату: спячка, миграции, состояние покоя в зимние месяцы. Почвы бурые и серые лесные. В этих районах человеческая цивилизация получила наибольшее развитие, поэтому большая часть широколиственных лесов заменена культурными сообществами.[ . ]
Листопадные леса умеренной зоны (широколиственные леса), расположенные южнее тайги, в отличие от нее, не имеют сплошного распространения (рис. 7.2). Произрастают они в условиях более мягкого климата, с осадками от 700 до 1500 мм/г, с умеренными температурами и четко выраженными сезонами. В основном в листопадных лесах среди древесной флоры доминируют бук и дуб. Благодаря опаду листьев формируется мощная лесная подстилка, позволяющая перезимовать многим беспозвоночным животным.[ . ]
Листопадные леса — это те районы суши, где человеческая цивилизация получила наибольшее развитие. Поэтому трудно сейчас найти широколиственные нетронутые леса. Большая их часть заменена культурными сообществами.[ . ]
Листопадные леса умеренных широт. Они произрастают в районах с невысокими средними температурами, значительно меняющимися по сезонам. Зимы здесь не очень суровы, летний период продолжителен, осадки выпадают равномерно в течение всего года. По сравнению с тропическими леса умеренного пояса быстро восстанавливаются после вырубки и, следовательно, более устойчивы к антропогенным нарушениям.[ . ]
Для листопадных лесов эти показатели соответственно равняются 1,6 и 1,0% для бо-реальных хвойных - 1,1 и 0,75%, для наиболее продуктивных природных травянистых сообществ - 0,6 и 0,5%, для площадей земледельческого использования 0,7 и 0,6%.[ . ]
Биом листопадных лесов Северной Америки подразделяется на много разновидностей, представляющих разные климаксные типы лесов. Вот некоторые из них.[ . ]
В зоне листопадных лесов вода может быть серьезным лимитирующим фактором только в конце лета, особенно в южной части этой зоны. Местная естественная растительность адаптирована к периодическим летним засухам, но некоторые выращиваемые здесь культуры к ним не адаптированы. Фермеры на юге США, которые приобрели горький опыт после неоднократной гибели урожая, начинают наконец переходить на орошение в конце лета. В районах выпадения зимних дождей основной продуктивный сезон— конец зимы и весна; в пустыне эффективный вегетационный сезон сильно сокращен.[ . ]
Сходные листопадные леса умеренного континентального климата распространены в Северной Америке, Европе и Восточной Азии. Злаковники умеренной зоны встречаются в условиях сходного климата в Северной Америке, Евразии и в Южном полушарии. Еще более широкие по объему группы конвергентных биомов, или формаций, различных континентов называются типами биомов, или типами формаций. Эти широкие категории так немногочисленны, что вполне могут быть описаны в этой книге. Концепция биомов и типов биомов обоснована для наземных сообществ. Ее можно распространить и на водные сообщества, но это представляется менее естественным, так как водные сообщества иначе переходят одно в другое и характеризуются менее четкими зависимостями структуры от климата.[ . ]
| Березовая роща в листопадных лесах умеренной зоны |  |
Биоценоз широколиственного леса. Среди наземных биоценозов наиболее сложным по структуре и разнообразным по видовому составу является лес. Леса занимают огромные пространства суши (около 26—30 %), это основной тип растительного покрова Земли. Они распространены на всех континентах, кроме Антарктиды. Это тропические леса Южной Америки, Африки и Азии, листопадные леса Европы и Северной Америки, тайга Сибири и Канады.[ . ]
Биомасса тропического дождевого леса вполне сравнима с биомассой лесов умеренного климата, содержание питательных веществ в биомассе того же порядка или несколько выше, чем в листопадном лесу умеренной зоны. Среднее содержание зольных элементов в тропических дождевых лесах составляет около 1,58%, содержание азота —0,57%. По сравнению с лесом умеренной зоны тропический дождевой лес продуцирует более обильный опад листьев, но он быстро разлагается, и поэтому количество подстилки на почве низкое (0,1—0,6 кг/м2). Количество органического вещества, дисперсно распыленного в глубоком выщелоченном слое в некоторых из этих почв, тем не менее велико и много выше, чем можно предполагать, судя по светлой окраске этих почв.[ . ]
Прекрасную сводку по растительности листопадных лесов читатель найдет в книге Брауна (1950).[ . ]
Он также характеризуется быстрым ростом и в благоприятных условиях достигает 50 м в высоту. Корневые подставки тетрамелеса очень сильно развиты и достигают иногда 6 м в высоту. Он встречается на острове Шри-Ланка, на западном побережье полуостровной Индии (включая Западине Гаты), в тропических муссонных лесах Сиккима и Ассама, в юго-восточной прибрежной части Бангладеша, на Андаманских остро- вах, на полуострове Малакка, на Малайском архипелаге (однако, отсутствует на Калимантане, Филиппинах и Молуккских островах), на Новой Гвинее и на полуострове Кейп-Йорк в Северо-Восточной Австралии. Тетрамелес произрастает в муссонных и особенно во влажных листопадных лесах и обычен в тикковых лесах. Обычно поднимается в горы не выше 500 м: над уровнем моря, но иногда до 1000 м.[ . ]
Растут симплоконые в подлеске вечнозеленых и листопадных лесов, в зарослях кустарников и на открытых пространствах. В Таиланде симплокосы образуют нижний ярус в вечнозеленых умеренных лесах. Встречаются они в бамбуковых и дубовых лесах, по краям мангровых зарослей, по берегам рек и потоков, по краям болот и на склонах холмов, нередко поднимаясь до высоты 2000 м над уровнем моря. В Восточных Гималаях известно их нахождение на высоте 3600 м над уровнем моря. Растут они на богатых алюминием почвах. Некоторые представители этого семейства красивы и культивируются как декоративные. Особенно привлекателен симплокос метельчатый, покрывающийся весной белыми душистыми цветками, а осенью украшенный кистями голубых плодов. Некоторое применение находят они и в медицине, и в красильной промышленности. Из корней симплокоса красильного (S. tincloria), обычного в подлеске густых североамериканских лесов, получают желтую краску. В Японии для этих целей используют листья симплокоса японского (S. lucida), а в Индии красную и желтую краску получают из коры и листьев симплокоса кистевидного (S. racemosa). Кору этого растения и некоторых близких видов индусы применяют в народной медицине.[ . ]
| Зависимость между чистой первичной продукцией листопадных лесов Северной Америки и длительностью вегетационного периода (Leith, 1975) |  |
Каликаптус и химонаитус произрастают главным образом в листопадных лесах и кустарниковых зарослях в умеренно теплом климате. Североамериканский род каликаптус включает один вид в Калифорнии и 3 вида в юго-вооточ-пых штатах США. Китайский род химоиаптус имеет 3 вида.[ . ]
Среди насекомых много вредителей, которые наносят большой вред лесам. Крупные животные в листопадных лесах, в принципе, те же, что и в тайге: лоси, медведи, рыси, лисицы и др., разнообразна и богата орнитофауна.[ . ]
| Профили трех типов почв. Слева |  |
Наибольшее видовое разнообразно их связано с тропическими вечнозелеными лесами. Они обитают в тенистых ущельях, па террасах рек и вблизи водопадов, на сырых скалах или растут на стволах деревьев в районах с частой облачностью, дождями и туманами. В Америке геснериевые встречаются преимущественно до 1200 м, реже до 2400 м (и до 3500 м) над уровнем моря. В Гималаях и в горах Китая они обычно поселяются па высоте от 2000 до 3000 м. Произрастают они и в горных листопадных лесах и кустарниковых зарослях, а и Африке (па восточных склонах Драконовых гор) н среди кустарииково-ксерофитпоп растительности. В Карибской и других флористических областях Америки в районах с наличием сухого и влажного сезонов года или в переменно-влажных лесах растения имеют ряд приспособлении для сохранения воды и питательных веществ.[ . ]
| Зависимость между прогнозируемой и наблюдаемой продукциями гетеротрофов в различных местообитаниях |  |
Так, биомом являются широколистные листопадные леса умеренной зоны в восточной части Соединенных Штатов. Прерии и прочие злаковники в условиях умеренно сухого климата Среднего Запада и Запада США — это тоже биом. Биом — это группа наземных экосистем данного континента, которые имеют сходную структуру или физиономию растительности и общий характер условий среды, что находит отражение в этой структуре и в характеристиках их животного населения.[ . ]
Вскоре после того, как стает снег, безлистные кусты их покрываются массой золотисто-желтых крупных колокольчатых цветков, оживляя яркими солнечными пятнами едва пробуждающуюся природу. Несколько позже начинается цветение сирени, без которой невозможно себе представить наши сады. Розовые, сиреневые, пурпурные, белые, голубоватые ее кусты наполняют сад топким ароматом и изысканной красотой. В культуре встречается до 20 видов сирени, по более всего распространена сирень обыкновенная (Syringa vulgaris), представленная в садах около 500 сортов. Дико сирень обыкновенная растет в горных областях в подлеске светлых листопадных лесов и на открытых местах в Южной Европе от Албании и Северо-Восточной Греции до Западных Карпат и доходит до Малой Азии. В Среднюю Европу сирень была введена только в XVT в. и долго оставалась там растением аристократических садов. Ныне этот неприхотливый кустарник встречается в культуре повсеместно, в СССР от западных до восточных границ и на север до Соловецких островов и Тобольска. В закрытом грунте растение может цвести зимой", поэтому сирень широко используют на выгонку.[ . ]
В зависимости от местности на каждый .квадратный метр земной поверхности ежеминутно падает от 0 до 5 Дж солнечной энергии. Если бы вся она фотосинтетически превращалась в биомассу (т. е. если бы к. п. д. фотосинтеза составлял 100%), растительного материала создавалось бы на порядок или два выше, чем в настоящее время. Однако значительная доля этой солнечной энергии растениям недоступна. В частности, по спектральному составу только около 44% падающего коротковолнового свега пригодно для фотосинтеза. Однако даже с учетом этого продукция будет гораздо ниже возможного максимума (см. также гл. 3). На рис. 17.7 в логарифмической шкале -представлена чистая эффективность фотосинтеза (процент перехода падающей фотосинтетически активной радиации, ФАР, в чистую первичную продукцию надземных частей растений). Эти данные получены в рамках МБП в США для семи хвойных, семи листопадных лесов и восьми сообществ пустыни. Наиболее высокая эффективность отмечена в хвойных лесах, но и там она составила только 1—3%. Листопадные леса способны превращать в биомассу 0,5—1 % ФАР, а пустыни, несмотря на более высокую освещенность,— лишь 0,01—0,02%. Максимальная эффективность фотосинтеза зерновых культур при идеальных условиях — 3—10% (табл. 17.2).[ . ]
В нескольких работах было показано, что структура пищевой цепи может влиять на ее упругость (быстроту возвращения к равновесному состоянию) в условиях колебаний притока энергии и биогенных элементов. О’Нил (O’Neill, 1976) рассматривал сообщество как трехкомпонентную систему, состоящую из активной растительной ткани (Р), гетеротрофных организмов (Я) и неактивного мертвого органического вещества (D) (рис. 21.10). Скорость изменения биомассы каждого из этих блоков зависит от переноса энергии между ними. Так, у Р она пополняется от одного источника (чистая первичная продукция) и теряется двумя путями (потребление гетеротрофами и переход к D в виде подстилки). Изменения Н определяются двумя способами как поступления (поглощение биомассы живых растений и мертвого органического вещества), так и потерь (дефекация и расходы на дыхание). Наконец, биомасса блока D также двумя путями пополняется (растительный спад, и дефекация) и расходуется (потребление гетеротрофами и физический вынос за пределы системы). Подставляя в расчеты реальные данные по шести сообществам, характеризующим-тундру, тропический лес, листопадный лес умеренного пояса, соленый марш, пресноводный ручей и пруд, О’Нил изучал на моделях этих сообществ стандартные нарушения (сокращение исходной биомассы на корню активной растительной ткани на 10%). Он следил за скоростями восстановления систем до равновесного состояния, сопоставляя их с притоком энергии на единицу биомассы живой ткани (рис. 21.11).[ . ]
Наши наблюдения над бентосом наводят на мысль, что растительные сообщества могли бы быть более разнообразными в условиях стабильной среды. Климат пустыни Сонора отчетливо неустойчив с чередованием в году двух влажных н двух сухих периодов и с высокой изменчивостью общего количества осадков в разные годы. Однако некоторые части пустыни Соиора весьма богаты видами (см. рис. 3-10). Оказывается, что в этой пустыне нестабильность природных условий не столько лимитирует разнообразие, сколько превращается в такой аспект среды, на который растения реагируют дифференциацией ниш (см. рис. 3-7) и, как следствие этого, разнообразием видов. Растительность Ближнего Востока очень значительно нарушена человеком, она подвергалась пожарам, рубкам, сильному и разнообразному давлению выпаса овец, коз, крупного рогатого скота и верблюдов. Но измененная выпасом структура редколесий и кустарников тем не менее очень богата видами, адаптировавшимися к этим нарушениям, особенно видами однолетних и луковичных растений. Тот факт, что растительные сообщества теплого климата в Соноре, на Ближнем Востоке и в других местах столь богаты видами, несмотря на засуху и нестабильность среды, наводит на мысль, что температура в большей мере, чем увлажнение или стабильность, является тем основным фактором, который определяет видовое разнообразие сосудистых растений. Можно упомянуть и еще одно наблюдение над сообществами наземных растений, а именно, широколиственные листопадные леса в среднем заметно богаче видами, чем вечнозеленые хвойные леса в сходных условиях среды. Тип доминантных видов, определяющих характер листового опада и химический состав органического вещества почвы, значительно влияет на видовое разнообразие наземных растительных сообществ.[ . ]
С разнообразием тропических типов климата, свойственных различным регионам, связана и смена основных типов биомов. Степень выраженности и продолжительность сухого периода, уменьшение суммы осадков, снижение влажности воздуха определяют изменение характера растительности и животного населения (рис. 61).
Растительность. В связи с уменьшением суммы осадков и возрастанием контрастности между влажным и сухим сезонами прослеживается ряд тропических лесных и кустарниковых сообществ, отличающихся по структуре, набору жизненных форм, сезонной ритмике, продуктивности. В этом ряду происходит постепенное снижение видового разнообразия.
При выраженности засушливого периода, продолжающегося от одного до двух с половиной месяцев, вечнозеленый тропический лес может существовать лишь при сумме годовых осадков не менее 2500 - 3000 мм. Однако и здесь наиболее высокие деревья сбрасывают листья одновременно, а эпифиты прекращают жизнедеятельность на сухое время.
При возрастании продолжительности засушливого периода или сокращении годовой суммы осадков меняется структура леса, наиболее высокорослыми оказываются листопадные породы, а под их пологом сохраняются вечнозеленые деревья. Такие леса называются полу вечнозелеными. При дальнейшем ухудшении условий увлажнения они сменяются листопадными тропическими лесами. Среди последних различаюттропические влажные листопадные леса (влажные муссонные леса) итропические сухие листопадные леса. К сухим листопадным лесам, к которым нередко относят и колючие, при дальнейшем возрастании сухости климата примыкаютредколесья исообщества колючих кустарников. Уменьшение годовой суммы осадков в сочетании с нарастанием продолжительности и степени выраженности сухого периода находит отражение в постепенном снижении видового разнообразия.
Тропические полувечнозеленые дождевые леса особенно хорошо развиты в муссонной области Индостана, Бирмы, Таиланда, п-ова Малакка, где непосредственно контактируют с дождевыми тропическими лесами, к которым они близки по структуре, отличаясь в то же время меньшим числом видов. В целом сохраняется тот же набор жизненных форм, разнообразие лиан и эпифитов. Различия проявляются именно в сезонной ритмике, в первую очередь на уровне верхнего яруса древостоя (до 30 % деревьев верхнего яруса относятся к листопадным видам). В то же время нижние ярусы включают большое число вечнозеленых видов. Травяной покров представлен главным образом папоротниками и двудольными.
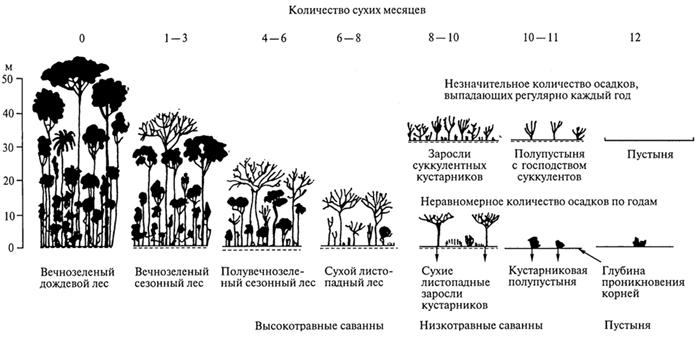
Рис. 61. Зависимость развития различных лесных формаций в тропических областях Перу от продолжительности сухого периода (Г.Вальтер, 1968)
В конце сухого периода многие деревья и лианы цветут, в то время как под ногами шуршат и пылят сухие листья, которые очень быстро разлагаются. В целом это переходные типы сообществ, местами в значительной степени сведенные человеком и замененные саваннами или плантациями.
Тропические влажные листопадные леса развиваются в таких гидротермических условиях, когда продолжительный сухой период (3 - 5 мес) сменяется очень влажным дождливым. Видовое разнообразие снижено, древостой верхнего яруса часто образован одним или двумя-тремя видами. Так, в Юго-Восточной Азии характерны леса с доминированием видов из родов диптерокарпус, саловое дерево, тик. В сезон дождей леса имеют вполне гигрофитный облик, лианы и эпифиты еще довольно многочисленны, однако видовой состав последних беден. Одна из важных структурных особенностей этих лесов - сомкнутый травяной покров из двудольных, папоротников с примесью геофитов (амариллисовые), а иногда и злаков. Цветение большинства видов сдвинуто на сухой период (рис. 62).
Если в самом общем виде смена от вечнозеленых тропических лесов к полулистопадным и влажным листопадным носит постепенный характер, то сухие листопадные леса, развивающиеся при
годовой сумме осадков 800-1000 мм, имеют ярко выраженный ксероморфный облик. Они хорошо различаются по упрощенной структуре, изменению набора жизненных форм и видового состава. Как правило, деревья невысоки (10-12 м или в некоторых типах леса 20 - 22 м). Характерны пирамидальные или уплощенные кроны, корявые, извилистые стволы; ветвление начинается невысоко над землей. Почки древесных пород защищены от засухи, стволы часто покрыты толстой коркой. Эпифиты почти отсутствуют, лиан мало. Так, в восточной части тропической Африки развит тип леса "миомбо", образованный видами из семейства бобовых. В вертикальной структуре выделяется кустарниковый ярус, часто обильны всходы и подрост деревянистых видов, травянистых растений немного. Однако при осветлении в связи с антропогенным воздействием сомкнутость травяного покрова возрастает и в нем увеличивается доля злаков (рис. 63).
Эти леса в течение многих тысячелетий подвергаются пожарам. Один из результатов их воздействия - формирование сложных сочетаний сухих лесов с облесенными саваннами и зарослями колючих кустарников.
Разнообразны тропические редколесья, часто сочетающиеся с сообществами колючих кустарников (рис. 64). Они встречаются в тех областях тропиков, где продолжительность засушливого периода составляет 8 - 9 мес, а сумма осадков варьирует от 350 - 800 мм. Это светлые труднопроходимые леса с высотой древесного полога 5- 12 м и сравнительно небольшим числом видов. Господствуют преимущественно колючие деревья, часто жестколистные, вечнозеленые, а также с листвой, опадающей на сухой период. Деревья обычно коренастые, с неправильным ветвлением, с кривыми ветвями; значительно реже встречаются деревья с прямыми стволами. Характерны деревья с суккулентными стволами, иногда

Рис. 63. Профильная диаграмма сухих листопадных (саванновых) лесов, льяносы Венесуэлы (G.Walter, S.-W.Breckle, 1985)
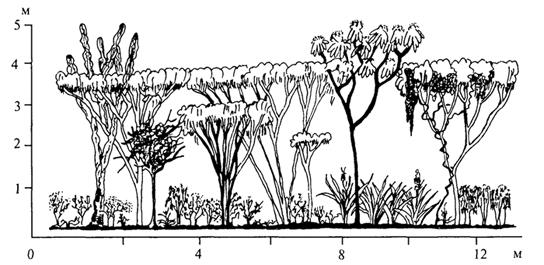
Рис. 64. Профильная диаграмма формаций колючих кустарников с преобладанием видов акаций, мимозы и других представителей семейства мимозовых в Венесуэле (G.Walter, S.-W. Breckle, 1985)
покрытыми толстой коркой, иногда гладкими, бутылкообразными или бочковидными. Сомкнутость крон очень небольшая. Здесь же растут и безлистные стеблевые суккуленты, канделябровидные и колоннообразные. Иногда именно стеблевые суккуленты определяют общий облик редколесий (рис. 65). Хорошо развит кустарниковый ярус, часто из колючих или безлистных прутьевидных растений. Много вьющихся растений (деревянистых или травянистых). Травяной ярус, как правило, выражен слабо.

Рис. 65. Профильная диаграмма суккулентных формаций Венесуэлы (G.Walter, S.-W. Breckle, 1985): 1-6 - стеблевые суккуленты; 7- 11 - кустарники; 12 - многолетние травы; 13, 14 - эпифиты
Животное население. Практически у всех видов животных наблюдается сезонная ритмика жизненных процессов. Это выражается в приуроченности периода размножения в основном к влажному сезону, в полном или частичном прекращении активности на время засухи у многих наземных и почвенных беспозвоночных, амфибий, в миграционных перемещениях на время неблагоприятного сухого сезона у некоторых насекомых, например саранчовых, птиц, рукокрылых и крупных копытных.
Общие запасы биомассы в различных вариантах листопадных лесов и редколесий существенно различаются. В лесах общая биомасса (сухая масса) составляет 200 - 250 т/га, а в редколесьях - не более 150 т/га. Еще меньше она для колючих кустарников - менее 100 т/га. Соответственно и годовая первичная продукция в этом ряду сообществ колеблется от 30 до 10 т/га в год. Запасы зоомассы составляют примерно 0,01 % общего объема биомассы.
риода — 8—9 месяцев, а сумма осадков — 350—800 мм. Это светлые труднопроходимые леса с высотой древесного полога 5—12 м и сравнительно небольшим числом видов растений.
Здесь господствуют колючие деревья, жестколистные, вечнозелёные, а также с листвой, опадающей на сухой период. Деревья обычно коренастые, с кривыми ветвями и стволами. Неповторимый облик тропическим редколесьям придают деревья с сукку-лентными (накапливающими влагу) бутылкообразными или бочковидны-ми стволами. Здесь же растут и безлистные суккуленты, похожие на колонны и канделябры.
Как и у растений, жизненные процессы почти у всех видов животных подчинены смене времён года. Так, размножаются они во влажный сезон, а многие беспозвоночные и амфибии на время засухи полностью или частично прекращают активность. С засухами связаны и миграции некоторых насекомых, например саранчи, а также птиц, рукокрылых и крупных копытных.

Тропический лес — тип биома, распространенный в экваториальном, субэкваториальном и тропических поясах Земли. Сложился в условиях избытка влаги и тепла. Занимает 5–6 % земной поверхности, в области своего распространения — 1/3 часть суши; площадь 2500–3000 млн. га. Площадь сомкнутых тропических лесов составляет: в Южн. и Центр. Америке свыше 600 млн. га, в Юго-Вост. Азии и Австралии — 300, в Африке — 216 млн. га.
В тропических лесах выделяют две основные группы формаций — дождевого, или влажного, тропического леса (гилея) и сезонного тропического леса (дождезеленого, зимнезеленого).
Дождезеленые тропические леса распространены в бассейне р. Амазонки, в Центр. Америке, на некоторых островах Карибского моря, на побережье Гвинейского залива, в бассейне р. Конго.
Тропические дождезеленые (сезонные) леса и редколесья Африки
«Арзамасский государственный педагогический
студентки V курса ЕГФ
Маркова Н., Шадрина М.

Географическая распространенность
Географическая распространенность
- В тропических лесах выделяют две основные группы формаций — дождевого, или влажного, тропического леса (гилея) и сезонного тропического леса (дождезеленого, зимнезеленого).
Дождезеленые тропические леса распространены в бассейне р. Амазонки, в Центр. Америке, на некоторых островах Карибского моря, на побережье Гвинейского залива, в бассейне р. Конго.


Ствол дерева покрыт эпифитными растениями разных видов: папоротниками, орхидеями и др.

Дерево фикуса со спускающимися воздушными корнями.

Гидрохимический режим почвы
Геохимический режим почвы
Сообщества фитоавтотрофов
- Дождевые тропические леса — наиболее богатая видами растительная формация на Земле (свыше 4/5 всех видов растений). Их флору считают центром эволюционной активности, откуда пополнялись все флоры мира, в том числе умеренных широт. Древесные виды в них явно преобладают (около 70 % высших растений), причем их видовое разнообразие исключительно высоко (от 40 до 100 на 1 га).
- Оно может проявляться только в наиболее засушливых районах , периодически заливаемых дождями и имеющих плохой дренаж из - за материнских пород тяжелого механического состава . В открытых саваннах почвы обеднены гумусом по сравнению с лесами , развитыми в тех же климатических условиях .
Экваториальный дождевой лес на востоке бассейна Конго (Заир).

В древостое тропических лесов доминируют представители семейств диптерокарповых, бобовых, миртовых, пальм и др., представлены также древовидные папоротники. В условиях экваториального пояса некоторые папоротники вырастают как деревья. Особенностью природной растительности является развитие лесов с очень густым древостоем и многоярусностью, незначительным травянистым покровом. Экваториальные леса вечнозеленые, с большим количеством видов деревьев. В таких лесах трудно найти рядом деревья одного вида.
В этих лесах очень много вьющихся растений — лиан и эпифитов, поселяющихся на других деревьях. Часто лианы, особенно на опушках, образуют непроходимые заросли. Высота деревьев во влажных экваториальных лесах достигает 40–60 м, толщина — до 4 м. Крона у них развита слабо, листья крупные, блестящие и толстые, а на стволах нередко образуются воздушные корни, которые обеспечивают дополнительное усвоение влаги и газов из воздуха. В тропических лесах деревья растут, плодоносят и меняют листву на протяжении всего года.

Строфант комбе
Строфант щетинистый
Пуэрария волосистая
Семейство логаниевые
- Это семейство включает немногим более 20 родов и около 500 видов, распространенных главным образом в тропических и субтропических странах, лишь немногие представители заходят в умеренные области.
Семейство кутровые

В структуре фитоценозов дождевого тропического леса обычно 3 яруса, которые редко выражены четко. В тропических лесах очень много болот с торфяниками, поросшими низкими кустарниками. Дождезеленые леса образованы главным образом невысокими ( около 10 — 12 м ) ветвистыми деревьями с перистыми листьями из семейства бобовых . Часто отчетливо доминируют немногие виды из родов брахистегия, изоберлиния , весьма обычны также некоторые виды акаций и альбицций .

Масличная пальма
Слоновая трава
Более засушливая полоса суданских саванн представлена главным образом регулярно выжигаемыми злаковниками , на фоне которых растут отдельные баобабы и группы небольших зонтиковидных деревьев и кустарников. Между суданской саванной и Сахарой протянулась переходная зона под названием Сахель , или сахельская саванна. Сахельская саванна представляет собой разреженное акациевое редколесье с очень скудным злаковым покровом из однолетних видов . На юге Африки также распространены разреженные акациевые саванны , переходящие в пустыню .

Африканский баобаб

Гетеротрофы первого порядка
В этих сообществах наличие термитов особенно заметно благодаря внушительным размерам их построек — термитников , высота которых нередко достигает 2 — 3 м , а в исключительных случаях — 9 метров и диаметр 30 м . В Восточной Африки обычно в одном небольшом районе саванн , редколесий или листопадных тропических лесов можно обнаружить 20 — 30 видов термитов , из которых 10 — 15 ведут скрытный , подземный образ жизни или обитают в стволах деревьев , а остальные сооружают заметные термитники . Среди всей конкретной фауны термитов относительно обильными оказываются лишь 5 — 6 видов , на которые приходится около 90% всего населения термитов.


Большинство термитов дождезеленых лесов и саванн относится к особым видам тех же родов , что представлены и в гилее . Это же справедливо и для большинства других сапрофагов. Отмечается лишь общее уменьшение видового разнообразия и появление сравнительно небольшого числа видов - доминантов среди разных групп организмов . В целом же такие группы , как нематоды , клещи - орибатиды , коллемболы , черви - мегасколециды , крупные кивсяки , различные тараканы , сверчки , уховертки весьма обильны и в дождезеленых лесах , и в саванновых редколесьях.
Читайте также: