
У бэтсон и р пеннет скрещивали растения душистого горошка
Обновлено: 19.09.2024
Свидетельство и скидка на обучение каждому участнику
Сцепленное наследование генов. Хромосомная теория наследственности
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Изучением наследования признаков не дающих независимого распределения генов занимался Томас Морган и его ученики. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Плодовая мушка является удобным объектом генетических исследований. Каждые две недели при температуре 25 °С мушка дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
опыты Моргана
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых).

При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями).
В — нормальные крылья
b — недоразвитые крылья
Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, соединены между собой. Оказалось, что гены образуют группы сцепления, т.е. гены одной группы наследуются сцеплено, а гены разных групп — независимо.
Все гены одной хромосомы образуют группу сцепления и наследуются совместно.
Количество групп сцепления соответствует гаплоидному набору хромосом. Поэтому Морган предположил, что гены локализованы на хромосомах.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме.
Группы сцепления разрушаются при кроссинговере, когда происходит обмен участками гомологичных хромосом в профазу I мейоза. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот.
Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb.
Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. Гибридные особи, имеющие иное сочетание признаков, чем у родителей называются рекомбинантами.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
хромосомная теория наследственности
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.

1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb , а отцовский — один тип — аb . Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb . Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb . Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В , появляются гаметы Аb и аВ , и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
| Образуются гаметы: |
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
| Образуются гаметы: |
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза).

1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
- гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- гены расположены в хромосомах в определенной линейной последовательности;
- гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- каждый вид имеет характерный только для него набор хромосом — кариотип.
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.

Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.

1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
 | Образуются гаметы: |  |
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
 | Образуются гаметы: |  |
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза).

1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Изучением наследования признаков не дающих независимого распределения генов занимался Томас Морган и его ученики. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Плодовая мушка является удобным объектом генетических исследований. Каждые две недели при температуре 25 °С мушка дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
опыты Моргана
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых).

При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями).
В — нормальные крылья
b — недоразвитые крылья

Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, соединены между собой. Оказалось, что гены образуют группы сцепления, т.е. гены одной группы наследуются сцеплено, а гены разных групп — независимо.
Все гены одной хромосомы образуют группу сцепления и наследуются совместно.
Количество групп сцепления соответствует гаплоидному набору хромосом. Поэтому Морган предположил, что гены локализованы на хромосомах.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме.
Группы сцепления разрушаются при кроссинговере, когда происходит обмен участками гомологичных хромосом в профазу I мейоза. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот.
Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb.
Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. Гибридные особи, имеющие иное сочетание признаков, чем у родителей называются рекомбинантами.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
хромосомная теория наследственности
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
- Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- Гены расположены в хромосомах в определенной линейной последовательности;
- Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- Сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- На основании частот рекомбинации определяют расстояние между генами. Что позволяет строить генетические карты хромосом.
ЗадачаУ кукурузы гладкая поверхность зёрен доминирует над морщинистой,неокрашенные зёрна рецессивный признак. Скрестили линию кукурузы с гладкими окрашенными зёрнами и с морщинистыми неокрашенными. В первом поколении все растения имели гладкие окрашенные зёрна. Гибридов первого поколения скрестили с родительским сортом с неокрашенными морщинистыми зёрнами. Получили 4152 растения с гладкими окрашенными зёрнами, 149 с морщинистыми окрашенными, 152 с гладкими неокрашенными и 4166 с морщинистыми неокрашенными. Как наследуются признаки. - РЕШЕНИЕ.
В первом поколении все растения с гладкими окрашенными зернами, след. эти признаки доминантны. Во втором поколении основными являются родительские фенотипы, значит гены сцеплены в транс-положении. Частота рекомбинантных фенотипов равна (149 + 152) : (4152+149+152+4166) = 301: 8619=0,035. Таким образом, расстояние между генами составляет 3,5 морганид.
Подавляющее большинство видов животных представлено особями двух полов — мужского и женского. Расщепление по половой принадлежности происходит в соотношении 1:1. Иными словами, у всех видов численность самцов и самок приблизительно одинакова. Еще Г. Мендель обратил внимание на то, что такое расщепление в потомстве по какому-либо признаку наблюдается в тех случаях, когда одна из родительских особей была гетерозиготой (Аа) по этому признаку, а вторая — рецессивной гомозиготой (аа). Было сделано предположение, что один из полов (тогда было неясно, какой именно) гетерозиготен, а второй гомозиготен по гену, который определяет пол организма. Современная теория наследования пола была разработана Т. Морганом и его сотрудниками в начале XX в. Им удалось установить, что самцы и самки различаются по набору хромосом.
У мужских и женских организмов все пары хромосом, кроме одной, одинаковы и называются аутосомами, а одна пара хромосом, называемых половыми, — у самцов и самок различается. Например, и у самцов, и у самок дрозофил в каждой клетке по три пары аутосом, а вот половые хромосомы различаются: у самок — по две Х-хромосомы, а у самцов X и Y (рис 1). Пол будущей особи определяется во время оплодотворения. Если сперматозоид содержит Х-хромосому, то из оплодотворенной яйцеклетки разовьется самка (XX), а если в сперматозоиде содержалась половая Y-хромосома — то самец (ХY). Соотношение полов при таком скрещивании теоретически всегда будет 1:1.
Так как у самок дрозофил образуются только яйцеклетки, содержащие половые Х-хромосомы, то женский пол у дрозофил называют гомогаметным. У самцов дрозофил образуются в равном соотношении сперматозоиды либо с Х-, либо Y-половыми хромосомами. Поэтому мужской пол у дрозофил называется гетерогаметным.
У многих видов живых существ, например у ракообразных, земноводных, рыб, большинства млекопитающих (в том числе и человека), женский пол гомогаметный (XX), а мужской — гетерогаметный (ХY).
Итак, у дрозофилы и человека женский пол является гомогаметным, и общая схема наследования пола у двух этих видов одинакова. У некоторых видов живых существ хромосомное определение пола совсем другое. Например, у птиц и рептилий — гомогаметны самцы (ZZ), а самки — гетерогаметны (ZW). У некоторых насекомых (например у сверчков) у самцов в хромосомном наборе лишь одна половая хромосома (ХО), а самки — гомогаметны (XX).
У пчел и муравьев половых хромосом нет, и самки имеют в клетках тела диплоидный набор хромосом, а самцы, развивающиеся партеногенетически (из неоплодотворенных яйцеклеток), — гаплоидный набор хромосом. Естественно, что в этом случае развитие сперматозоидов у самцов идет без мейоза, так как уменьшить число хромосом менее гаплоидного набора невозможно.
У крокодилов половые хромосомы не обнаружены. Пол зародыша, развивающегося в яйце, зависит от температуры окружающей среды: при высоких температурах развивается больше самок, а в том случае, если прохладно, — больше самцов.

Сцепленное наследование

Три к одному – Такой закон! Как много дум Наводит он…

Задача Грегор Мендель

Опыты У. Бетсона и Р. Пеннета В 1906 году английские генетики В. Бетсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружил, что эти признаки не дают независимого распределения в потомстве. Потомки всегда повторяли признаки родительских форм.
Генотип человека – тысячи различных признаков – размещается всего в 46 хромосомах. Это означает, что каждая хромосома содержит множество генов. Законы Менделя справедливы только для генов, локализованных в разных хромосомах.

Томас Морган Американский биолог и генетик. Лауреат Нобелевской премии.

ØМушка каждые две недели при температуре 25 С даёт многочисленное потомство. ØСамец и самка внешне хорошо различимы – у самца брюшко меньше и темнее. ØОни имеют отличия по многочисленным признакам. ØМушки могут размножаться в пробирках на дешёвой питательной среде Объект исследования Моргана – плодовая мушка дрозофила ØНаличие всего 4 -х пар хромосом

Закономерности сцепленного наследования признаков

Расщепление по фенотипу при независимом и сцепленном наследовании признаков

Группы сцепления Сцепленные гены – гены, Число групп сцепления равно расположенные в одной числу хромосом в гаплоидном хромосоме и наследуемые наборе: совместно. Дрозофила – 4 Горох – 7 Количество генов в различных Кукуруза – 10 группах сцепления (то есть в Мышь – 20 различных хромосомах) может отличаться друг от друга. Человек – 23 Дождевой червь – 36 Ящерица – 38 Кролик – 44 Корова -60
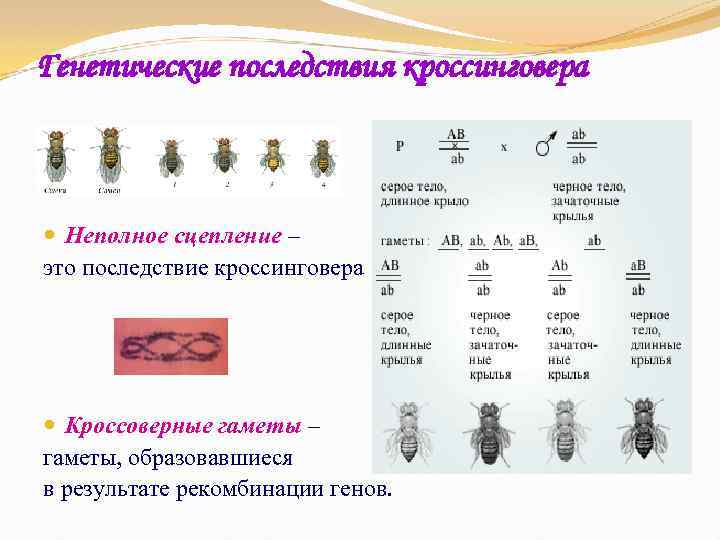
Генетические последствия кроссинговера Неполное сцепление – это последствие кроссинговера Кроссоверные гаметы – гаметы, образовавшиеся в результате рекомбинации генов.
Видео Конъюгация и кроссинговер

Кроссинговер Видео Конъюгация и кроссинговер

Хромосомная теория Гены располагаются в хромосомах в определенной линейной последовательности. В хромосоме каждый ген занимает определенное место (локус). Гены, расположенные в одной хромосоме, наследуются совместно, сцепленно.

Хромосомная теория Каждый вид имеет определенное количество групп сцепления, соответствующее числу хромосом в гаплоидном наборе. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом.
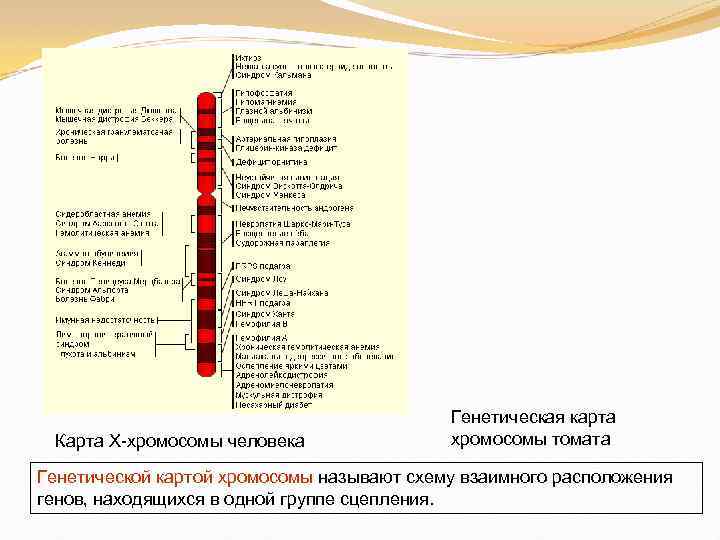
Генетическая карта Карта X-хромосомы человека хромосомы томата Генетической картой хромосомы называют схему взаимного расположения генов, находящихся в одной группе сцепления.
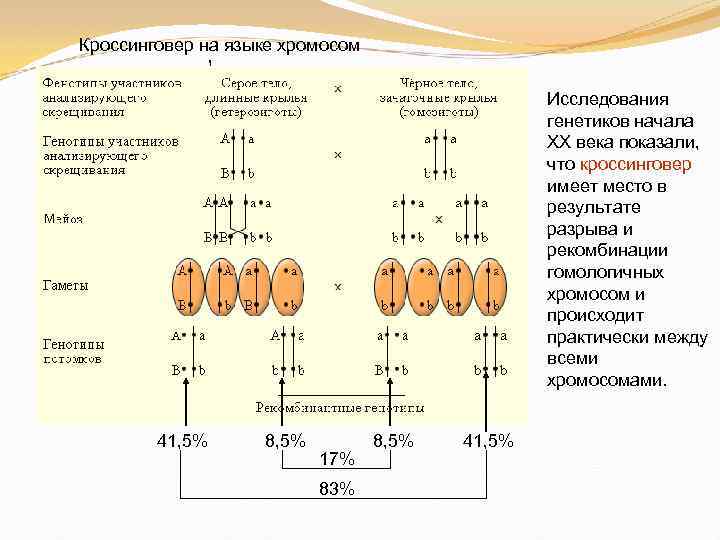
Кроссинговер на языке хромосом Исследования генетиков начала XX века показали, что кроссинговер имеет место в результате разрыва и рекомбинации гомологичных хромосом и происходит практически между всеми хромосомами. 41, 5% 8, 5% 41, 5% 17% 83%
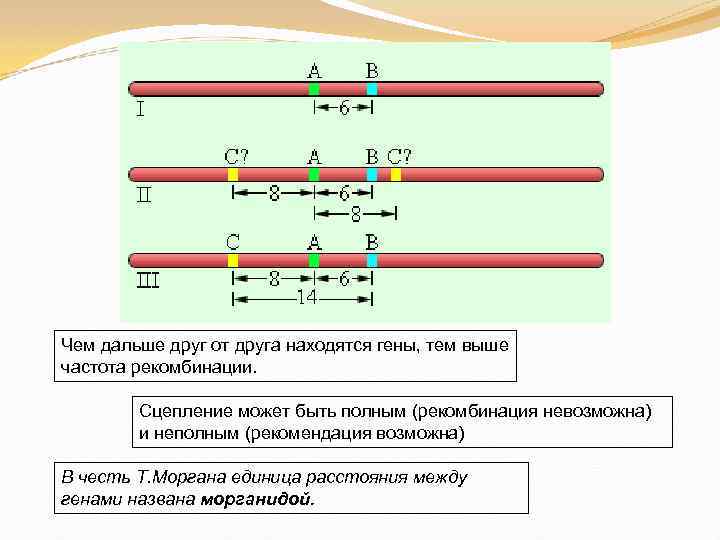
Чем дальше друг от друга находятся гены, тем выше частота рекомбинации. Сцепление может быть полным (рекомбинация невозможна) и неполным (рекомендация возможна) В честь Т. Моргана единица расстояния между генами названа морганидой.

Решение генетических задач На результаты количественного и качественного учета расщепления по генотипу и фенотипу Задача: Скрещены две породы кроликов: пятнистые нормальношерстные и сплошь окрашенные ангорские. В F 1 все кролики пятнистые нормальношерстные. В результате анализирующего скрещивания получено 26 пятнистых ангорских, 144 сплошь окрашенных ангорских, 157 пятнистых с нормальной шерстью и 23 сплошь окрашенных с нормальной шерстью. Вопросы: 1. Сколько разных фенотипов получится от такого скрещивания? 2. Какой процент гамет самки содержит одновременно гены пятнистой и ангорской шерсти? 3. Какой процент гамет содержит гены сплошной окраски и нормальной длины? 4. Какой процент потомков от этого скрещивания будет фенотипически похож на отца? 5. Какой процент потомков от этого скрещивания будет дигетерозиготен?

Дано: А – ген пятнистой окраски а – ген сплошной окраски В – ген нормальной длины в – ген ангорской шерсти - генотип самки ( по некроссоверным потомкам, которых более 50%) - генотип самца (т. к. скрещивание анализирующее) Ответ: Р × 1. 4 фенотипа 2. 7, 4 % (26÷ 350∙ 100) Г 3. 6, 6% (23÷ 350∙ 100) 4. 41, 2% (144÷ 350∙ 100) 5. 44, 2% (157÷ 350∙ 100) F 1 26 144 157 23

Решение задач: У кроликов пятнистость доминирует над сплошной окраской, а нормальная шерсть над ангорской. (Имеет место кроссинговер. ) Скрестили чистые линии породы ангорских пятнистых кроликов с породой, имеющей сплошь окрашенную нормальную шерсть. Гибриды F 1 были скрещены со сплошь окрашенными ангорскими кроликами. Определите, какие генотипы и фенотипы получатся в результате второго скрещивания?

СПАСИБО ЗА УРОК!
Читайте также: