
В пыльцевых зернах созревают
Обновлено: 18.09.2024
Палинологический анализ основан на различиях морфологической структуры пыльцевых зёрен и спор разных видов растений. Важными характеристиками для анализа являются размеры и форма пыльцевых зёрен и спор, типы апертур и их количество, виды скульптуры и текстуры [Нокс, 1985; Бурмистров, Никитина 1990; Мейер-Меликян, 1999; Halbritter, 2009].
Оболочка пыльцевых зёрен и спор – спородерма – представляет собой совокупность морфологически различных слоев: перина (периспорий), экзина (экзоспорий) и интина (эндоспорий), которые защищают цитоплазму пыльцевого зерна (споры) от физических, химических, микробиологических воздействий, обеспечивают дыхание, водообмен и другие функции (рис. 1, 2).
Рис. 1. Строение пыльцевого зерна [по Мейер-Меликян, 1999]
1 – эктэкзина, 2 – интина, 3 – эндэкзина, 4 – ядро вегетативной клетки, 5 – вегетативная клетка, 6 – апертура, 7 – генеративная клетка, 8 – ядро генеративной клетки
Рис. 2. Строение оболочки пыльцевого зерна [по Куприяновой, Алёшиной, 1972]
1 – элементы скульптуры, 2 – покровный слой, 3 – столби-
ковый слой, 4 – подстилающий слой, 5 - наружный слой, 6 - внутренний слой
Структура слоёв спородермы весьма специфична для порядков, семейств, а нередко родов и видов высших растений. Наружний слой (периспорий) характерен преимущественно для спор папоротников и мхов. У пыльцевых зёрен семенных растений перина встречается крайне редко: у некоторых голосеменных на ее месте формируется орбикулярный слой, а у покрытосеменных её заменяет трифина, особенности состава и строения которой определяются способом опыления [Мейер-Меликян, 1999].
Экзина, наружная часть оболочки пыльцевого зерна, подразделяется на два основных слоя: наружный (эктэкзина, сэкзина) и внутренний (эндэкзина, нэкзина). Между ними иногда может присутствовать промежуточный слой (мэкзина), который интенсивно окрашивается фуксином. Каждый из указанных слоёв имеет специфическое строение.
Эктэкзина большей частью представлена столбиками, которые являются ее основным элементом. Если они соединяются, то образуется покров. У некоторых видов зерен имеется надпокров (шипы, бугорки и т.д.). Соединённые ножки колонок образуют подстилающий слой (рис. 2). Эндэкзина – это обычно тонкий, почти незаметный слой, который не окрашивается фуксином.
Интина, внутренняя часть оболочки пыльцы, состоит из двух слоев: наружного (экзинтина, гиалины), образующего у пыльцы многих двудольных растений онкусы (линзовидные утолщения), и внутреннего слоя (собственно интины, эуинтины) [Куприянова, Алёшина, 1972].
Обычно 4 пыльцевых зерна (тетрада), возникшие в результате мейоза одной материнской клетки микроспор (рис. 3), при созревании становятся полностью свободными и в дальнейшем существуют в виде монад (одиночные), но некоторые и после созревания остаются в виде диад, тетрад, полаид или в поллиниях (рис. 4) [Мейер-Меликян, 1999].
Рис. 3. Фото тетрады, образовавшейся в резуль-
тате мейоза [Handbook of support for laboratory technicians (Atmosnet)]
Рис. 4. Одиночные и собранные пыльцевые зёрна, типы тетрад [по Куприяновой, Алёшиной, 1972]
А – монада, Б – диада, В, Г – тетраэдрическая, Д – квадратная, Е – крестообразная, Ж – ромбическая, З – линейная, И – полиады, К – поллиний
На стадии тетрад закладываются основные морфологические признаки пыльцевых зёрен, в частности определяется их полярность (рис. 5).
Часть поверхности пыльцевого зерна, обращённая к центру тетрады, называется проксимальной, противоположная ей поверхность, обращённая наружу и максимально удаленная от тетрады, называется дистальной. В соответствии с этим, у каждого пыльцевого зерна имеется 2 полюса – дистальный и проксимальный. Точка, расположенная в центре дистальной поверхности, называется дистальным полюсом. Точка, расположенная в центре проксимальной поверхности называется проксимальным полюсом [Wodehouse, 1935; цит. по Дзюба, 2005].
Пыльцевые зёрна с хорошо выраженными полюсами, морфологически неотличимыми друг от друга, называются равнополярными (изополярными), морфологически отличающимися друг от друга полюсами – разнополярными (гетерополярными), с невыраженными полюсами – неполярными (аполярными) (рис. 6).
Рис. 5. Полярность пыльцевых зёрен в тетраде [Handbook of support for laboratory technicians (Atmosnet)]
А – дистальный полюс, Б – проксимальный полюс, P – полярная ось, E – экваториальный диаметр
Рис. 6. Виды пыльцевых зёрен с различной полярностью (фото Р.Г. Курманова)
А – изополярное пыльцевое зерно (Vicia cracca – горошек мышиный), Б – гетерополярное пыльцевое зерно (Pinus sylvestris – сосна обыкновенная), В – аполярное пыльцевое зерно (Dianthus acicularis – гвоздика иглолистная)
Воображаемая линия, соединяющая проксимальный и дистальный полюса, называется полярной осью (Р). Экваториальной осью или диаметром (Е) называется линия, расположенная на экваториальной плоскости перпендикулярно полярной оси (рис. 5). Форма пыльцевых зёрен зависит от соотношения длины полярной оси к экваториальному диаметру (P/E) (табл. 1).
пылинка, мужской Гаметофит семенного растения; начинает развитие из микроспоры (См. Микроспора) в микроспорангии и завершает его после опыления, т. е. перенесения в пыльцевую камеру семяпочки (у голосеменных) или на рыльце пестика (у покрытосеменных растений). П. з. имеет 2 оболочки: наружную — экзину, прочную и стойкую, и внутреннюю — интину, состоящую главным образом из клетчатки и пектиновых веществ. В экзине обычно имеются тонкие участки или отверстия, т. н. апертуры — борозды или поры, через которые при прорастании П. з. выпячивается протопласт, покрытый интиной, т. е. происходит образование пыльцевых трубок (См. Пыльцевая трубка).
У голосеменных П. з. ко времени опыления состоит из нескольких живых клеток (у некоторых — и из остатков отмерших), среди которых имеются вегетативная (гаусториальная) и генеративная (антеридиальная); первая из них в пыльцевой камере образует пыльцевую трубку (гаусторий), внедряющуюся в Нуцеллус, вторая, делясь, — сперматогенную, или спермиогенную, клетку и сестренскую клетку (т. н. ножку). Сперматогенная клетка образует затем Гаметы — многожгутиковые сперматозоиды (например, у саговников) или безжгутиковые спермии (например, у хвойных), которые доходят до архегониев женского заростка по пыльцевым трубкам. У покрытосеменных ко времени попадания на рыльце пестика П. з. состоит либо из сифоногенной клетки и находящейся внутри неё спермиогенной клетки (двуклеточная или двуядерная пыльца), либо, если спермиогенная клетка уже разделилась, — из 2 спермиев, находящихся внутри сифоногенной клетки (трёхклеточная или трёхъядерная пыльца). При прорастании трёхклеточного П. з. в пыльцевую трубку попадают ядро сифоногенной клетки и оба спермия; при прорастании двуклеточного — ядро сифоногенной клетки и спермиогенная клетка, которая делится на 2 спермия в пыльцевой трубке. Развитие П. з. завершается по достижении пыльцевой трубкой зародышевого мешка (См. Зародышевый мешок), в который и попадают оба спермия, участвующие в двойном оплодотворении (См. Двойное оплодотворение).
У большинства семенных растений П. з. одиночные (монады); у некоторых покрытосеменных микроспоры и развивающиеся из них П. з. соединены по 2 (диады; например, у шейхцерии), по 4 (тетрады; у многих вересковых, некоторых орхидных и др.), по 8—12—16—32 (полиады; у мимозовых); у ластовневых, некоторых орхидных соединёнными остаются все П. з. одного или двух гнёзд пыльника (т. н. поллинии (См. Поллиний)).
Форма, размеры, строение П. з., особенно строение его экзины, скульптура поверхности, строение апертур и их положение разнообразны, но постоянны у растений одного вида; а у представителей различных таксонов, как правило, тем более сходны, чем ближе их родство. Поэтому изучение П. з. важно для систематики растений (см. Палинология). В связи со стойкостью экзины, хорошо сохраняющейся в осадочных породах, изучение П. з. лежит в основе одного из методов палеоботанического исследования — спорово-пыльцевого анализа (См. Спорово-пыльцевой анализ).
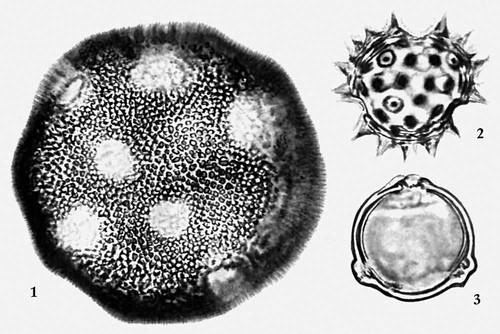
Пыльцевые зёрна: 1 — многопоровое насекомоопыляемого растения повоя заборного; 2 — трехбороздное насекомоопыляемого растения белокопытника холодного; 3 — трехпоровое ветроопыляемого растения грабинника.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Строение пыльцевого зерна
Функционально пыльцевое зерно представляет собой биологический контейнер, в котором содержится мужской гаметофит покрытосеменных и голосеменных растений [55]. Подобный контейнер защищает мужские гаметы от неблагоприятных воздействиий окружающей среды во время их передачи c пыльников на пестик.
Пыльцевые зерна различных видов растений варьирует как по размеру (от 10 до почти 100 мкм) так и по форме: круглые, овальнные, дисковидные, фасолевидные, а также нитевидные. Естественный цвет пыльцы в основном белый, кремовый, желтый или оранжевый. По текстуре клеточной стенки пыльца также различна, от гладкой до покрытой различными выростами.
Оболочка пыльцевого зерна состоит из двух главных слоев - экзины и интины (рис. 2). Строение этих двух слоев, так же как и их толщина – важные таксономические признаки растений.
Интина представляет собой внутренний слой оболочки пыльцевого зерна. Она облегает его содержимое и служит материалом, образующим пыльцевую трубку. Ее химический состав неоднороден: наружный слой образован преимущественно пектином, в состав же внутреннего слоя входят, главным образом, целлюлоза и пектин. Интина легко разрушается под действием кислот и щелочей и в ископаемом состоянии не сохраняется. Интина имеется у пыльцевых зерен всех цветковых растений.
Экзина — это внешний слой оболочки пыльцевого зерна. Благодаря основному ее компоненту — спорополленину, экзина характеризуется необычайной стойкостью: в отличие от интины она не растворяется в кислотах и щелочах, выдерживает температуру до 100°С и сохраняется миллионы лет в геологических отложениях. Спорополленин защищает вегетативную и генеративную клетки пыльцевых зерен от механических повреждений, химического развала, высыхания и ультрафиолетового излучения. На поверхности наружной стенки можно найти иногда зерна покрытые жидкостью, жирового вещества, так называемые "pollenkit".
Экзина состоит из двух основных слоев — эктэкзины и эндэкзины, к которым у многих растений прибавляется средний слой — мезэкзина.
Эктэкзина (ectexina, от греч. ektos — вне, снаружи), или внешняя экзина, представляет собой наиболее устойчивую часть экзины и отличается исключительным разнообразием строения. У цветковых растений различают 3 основных типа эктэкзины (рис. 3): столбиковую, или колумеллятную (ее называют также бакулатной), гранулярную и гомогенную.

Рисунок 2. Строение пыльцевого зерна [56]. 1 – эктэкзина, 2 – интина, 3 – эндэкзина, 4 – ядро вегетативной клетки, 5 – вегетативная клетка, 6 – апертура, 7 – генеративная клетка, 8 – ядро генеративной клетки
Гораздо менее характерна для цветковых растений гранулярная эктэкзина. Она бывает с тектумом или чаще без него, с подстилающим слоем или без него.
Очень редко встречается гомогенная эктэкзина, характерная для некоторых примитивных родов, как дегенерия и др. Наряду со структурой эктэкзины, т. е. Ее внутренним строением, очень важен также характер скульптeры ее поверхности. Различают многочисленные типы поверхностных скульптур. Многие скульптурные элементы настолько малы (меньше 0,1 мкм), что могут быть обнаружены только с помощью электронного микроскопа (рис. 4). Следует отметить, что аллергенные свойства оболочки пыльцевого зерна практически не исследованы.
Наряду со структурой эктэкзины, т. е. ее внутренним строением, важен также характер скульптуры ее поверхности. Различают многочисленные типы поверхностных скульптур. Многие скульптурные элементы настолько малы (меньше 0,1 мкм), что могут быть обнаружены только с помощью электронного микроскопа (рис. 4).
Наряду со структурой эктэкзины, т. е. ее внутренним строением, важен также характер скульптуры ее поверхности. Различают многочисленные типы поверхностных скульптур. Многие скульптурные элементы настолько малы (меньше 0,1 мкм), что могут быть обнаружены только с помощью электронного микроскопа (рис. 4).
Эндэкзина (endexina, от греч. endon — внутри), или внутренняя экзина, обычно гораздо тоньше эктэкзины, часто очень тонкая и нередко даже отсутствует. Как и эктэкзина, она устойчива к действию кислот и щелочей, но в отличие от нее не окрашивается основным фуксином. В то время как эктэкзина состоит в основном из радиальных структурных элементов, структура эндэкзины пластинчатая (ламеллятная), зернистая (гранулярная) или часто гомогенная.
Как правило, в экзине пыльцевого зерна имеются эластичные, гибкие, чаще тонкие или даже перфорированные места, служащие для выхода пыльцевой трубки- апертуры. Строение апертур, их число и расположение на поверхности пыльцевого зерна являются важнейшими признаками при установлении морфологических типов пыльцы.
Пыльца часто бывает собрана в диады (шейхцерия), тетрады (рогоз), поллиады (мимозовые), поллинии (орхидные) (рис. 5).
Укреплению тетрад или сцеплению отдельных пыльцевых зерен способствуют также висциновые нити, прикрепленные к проксимальному полюсу зерен или вблизи него. Длина нити достигает 1500 мкм. Висциновые нити характерны для пыльцы немногих семейств насекомоопыляемых растений: вересковых, кипрейных, протейных, некоторых нимфейных [57]. Строение пыльцевых зёрен несомненно имеет большое значение для проявления или отсутствия аллергенных свойств у пыльцы разных видов растений, однако изучено в данном аспекте к настоящему времени очень мало.
Актуальным остается поиск новых аллергенов, в частности, активно ведутся работы по изучению аллергенного состава поверхности пыльцевых зерен. Полагают, что поверхностные составляющие пыльцы вносят свой вклад в развитие аллергенных заболеваний [59, 60], но в целом, аллергенные свойства пыльцевой поверхности мало изучены. Главным образом, это связано с тем, что при получении пыльцевые экстракты проходят стадию промывки органическими растворителями с целью избавления от возможной контаминации загрязняющими веществами и микроорганизмами [61]. В результате, с зерен пыльцы смываются поверхностные молекулы внутреннего слоя целлюлозы (интина), внешней стенки (экзины), и внеклеточного матрикса, содержащего липофильные белки и липиды [62]. Поэтому при идентификации пыльцевых аллергенов растворимых белковых экстрактах поверхностные составляющие пыльцевых зерен остаются незамеченными.
Vigh-Conrad К.А. et al. анализировали уровень sIgE в различных сыворотках в микрочиповом формате и сравнивали аллергенные свойства различных пыльцевых фракций (поверхностной, цитоплазматической фракций и коммерческих препаратов пыльцы) [63]. Микрочипы содержали поверхностные и коммерческие экстракты пыльцы 22 видов растений, коммерческие экстракты 9 непыльцевых аллергенов и 5 рекомбинантных аллергенных белка. Для разделения поверхностной и цитоплазматической фракций образцы экстрагировались органическими растворителями или водными буферами, соответственно. Аллергены в микрочипах инкубировали с исследуемой сывороткой, после чего уровень sIgE оценивали путем непрямой иммунофлюоресценции. Скрининг продемонстрировал, что подобный микрочиповый анализ хорошо отражает уровень sIgE в сыворотке. Было обнаружено различие в аллергенном составе поверхностных и цитоплазматических пыльцевых фракций.

Рисунок 3. Схема строения оболочки пыльцевых зерен с двумя типами эктэкзины (столбиковой и гранулярной) [57].

Рисунок 4. Скульптура экзины пыльцевых зерен некоторых цветковых растений под сканирующим электронным микроскопом: 1 - морщинистая у купальницы китайской (Trollius chinensis, увел. x12600); 2 - мелко-остробугорчатая у нотофагуса клиффортиевидного (Nothofagus cliffortioides, увел. x3000); 3 - гладкосетчатая у морозника абхазского (Helleborus abchasicus, увел. x10000); 4 - сетчато-бугорчатая у гедиосмума ветвистого (Hedyosmum racemosum, увел. x10000); 5 - разнобугорчатая у скополии тангутской (Scopolia tangutica, увел. x6500); 6 - струйчатая у розы коричной (Rosa cinnamomea, увел. x6000); 7 - крупношиповатая у лапагерии розовой (Lapageria rosea, увел. x6000); 8 - сетчато-крупнобугорчатая у лилии Кесселринга (Lilium kesselringianum, увел. x5000); 9 - струйчато-бугорчато-дырчатая у скополии китайской (Scopolia sinensis, увел. x6500) [57].

Рисунок 5. Одиночные и собранные пыльцевые зёрна, типы тетрад [58]. А – монада, Б – диада, В, Г – тетраэдрическая, Д – квадратная, Е – крестообразная, Ж – ромбическая, З – линейная, И – полиады, К – поллиний.
ПЫЛЬЦЕВОЕ ЗЕРНО — мужской гаметофит семенных растений, который развивается из микроспоры в микроспорангии. Зрелое пыльцевое зерно покрыто спородермой, состоящей из экзины и интины. Экзина – многослойная оболочка, имеющая сложный химический состав, очень прочная, снаружи гладкая (например, у злаков) или структурированная (сложноцветные). В экзине имеются поры, через которые происходит прорастание пыльцы. Интина — тонкая, полупроницаемая пленка. Зрелый мужской гаметофит покрытосеменных растений является трехклеточным и состоит из вегетативной клетки и двух спермиев, которые образуются в результате деления генеративного ядра. Это деление может происходить либо внутри пыльцевого зерна в пыльнике, либо в пыльцевой трубке при прорастании пыльцы на рыльце. У голосеменных пыльцевое зерно содержит несколько вегетативных клеток и два спермия.
Читайте также: