Созревание тычинок в цветке
Обновлено: 18.09.2024
Морфологически и функционально тычинка соответствует микроспорофиллу. Существенной частью тычинки является микроспорангий — пыльник, в котором образуются микроспоры — пыльца.
Тычинка состоит из тычиночной нити, связника и пыльников. Обычно образуется два пыльника, в каждом из которых имеется два пыльцевых гнезда. Однако в некоторых случаях пыльцевых гнезд может быть меньше или больше. Связник — осевая часть тычинки, которая как бы является продолжением тычиночной нити и соединяет между собой пыльники.
Хорошо развитые тычинки не обнаруживают внешнего сходства с листом. Но это сходство проявляется в некоторых чертах анатомического строения и в процессе онтогенеза. Некоторые примитивные тычинки имеют расширенную пластинчатую нить, отдаленно напоминающую лист. В этом случае увеличивается и анатомическое сходство с листом.
Тычинки обнаруживают большое разнообразие формы, что обусловлено способом прикрепления пыльников и характером их раскрывания.
Строение тычиночной нити очень простое. Снаружи она покрыта эпидермисом, клетки которого слегка вытянуты, имеют кутикулу. Устьица на тычиночной нити обычно отсутствуют. Под эпидермисом располагается мезофилл, состоящий из немногих округлых тонкостенных клеток. Центральное положение в тычиночной нити занимает проводящий пучок.
Пыльник имеет многослойные стенки, ограничивающие пыльцевое гнездо. Наружным слоем служит эпидермис, состоящий из мелких плоских клеток с хорошо развитой кутикулой. Под эпидермисом располагается фиброзный слой. Клетки его характеризуются неравномерным утолщением стенок. Размеры, форма клеток этого слоя, характер утолщений в клеточных оболочках различны у разных растений. Фиброзный слой способствует раскрыванию пыльника. У некоторых растений он отсутствует.
Самый внутренний слой в стенках пыльника — это выстилающий слой, или тапетум. Он непосредственно выстилает пыльцевое гнездо и участвует в питании созревающей пыльцы. Клетки тапетума относительно крупные, тонкостенные, богатые зернистой цитоплазмой. Они часто бывают многоядерными или содержат полиплоидные ядра. У большинства растений тапетум — недолговечная ткань. Ко времени созревания спор клетки тапетума разрушаются. Цитоплазма их сливается, образуя тапетальный плазмодий, который распространяется в пыльцевом гнезде. Продукты растворения тапетума содержат много Сахаров и жиров, смешанных со слизистыми веществами.
Пыльцевое гнездо заполнено в зависимости от степени развития пыльника пыльцой или материнскими клетками пыльцы, или тканью, предшествующей развитию материнских клеток, — археспорием.
Связник состоит из паренхимной ткани и имеет проводящий пучок, который является продолжением проводящего пучка тычиночной нити. Иногда проводящий пучок в связнике ветвится. У разных растений связник развит в разной степени.
Развитие тычинки начинается с образования на конусе нарастания небольшого бугорка. В центре его обособляются крупные, богатые цитоплазмой клетки — это материнские, или первичные, клетки археспория. В результате деления первичных клеток археспория тангентальными перегородками обособляется тапетум, образующийся из наружного слоя клеток. Из внутреннего слоя возникают вторичные клетки археспория, или материнские клетки пыльцы.
Пыльца образуется из вторичных клеток археспория путем их редукционного деления (мейоза). Каждая материнская клетка дает четыре гаплоидные — тетраду спор. Это и есть клетки пыльцы, или пыльцевые зерна. Каждое пыльцевое зерно представляет собой гаплоидную клетку с двухслойной оболочкой. Наружный слой оболочки называется экзиной, внутренний — интиной. Экзина более плотная, имеет часто местные утолщения, которые придают пыльцевой поверхности характерный вид. В экзине имеются поры. Интина, более тонкая и мягкая часть оболочки, состоит из пектина.
Тычи́нка (лат. Stamen, устар. пыльцелистик[1]) — часть цветкацветковых растений, мужской репродуктивный орган, в котором образуется пыльца.
Строение тычинки
Тычинка состоит из тычиночной нити (стерильной части) и пыльника, в котором образуются пыльцевые зёрна. Тычиночная нить является гомологоммикроспорофилла разноспоровых плауновидных и голосеменных[2].
В каждом из четырёх гнёзд пыльника из клеток археспория после мейоза образуются микроспоры, которые затем прорастают в мужские гаметофиты — пыльцевые зёрна. Совокупность пыльцевых зёрен называется пыльцой[2].
Число тычинок в одном цветке может быть от одной до нескольких тысяч (например, у некоторых кактусов). Совокупность тычинок цветка называется андроцеем. Тычинки обычно расположены на цветоложе спирально или по кругу.
Тычинки могут полностью или частично срастаться.
У тыквенных они срастаются полностью, у астровых (сложноцветных) — пыльниками, у бобовых — тычиночными нитями (сросшиеся тычиночные нити в цветках представителей бобовых называются тычиночными трубками[2]).
У раффлезиевых и ряда представителей орхидных тычинки срастаются с пестиком, образуя особый орган, называемый колонкой.
Каждая тычинка в типичном случае состоит из двух основных частей: стерильной — тычиночной нити, или филаментума, и фертильной — пыльника, или микроспорангия.
Тычиночная нить у некоторых растений может отсутствовать и пыльники таких тычинок называются сидячими.
У некоторых растений тычиночные нити срастаются между собой, как, например, у зверобойных, бобовых, у других — ветвятся, как у клещевины.
Эпидермис тычиночной нити состоит из удлиненных по оси клеток с тонкой кутикулой, а нередко и совсем без кутикулы. Клетки эпидермиса тычиночной нити соединены между собой не столь плотно, как клетки эпидермиса листьев.
Устьица на тычиночных нитях встречаются иногда в значительном числе. Мезофилл нормальной тычиночной нити состоит из паренхимных клеток с тонкими оболочками и хорошо выраженными межклетниками. Многочисленные и обширные межклетники и тонкость оболочек клеток придают мягкость и рыхлость структуре-мезофилла тычиночных нитей.
В центре типичной тычиночной нити расположена сосудистая система первичного строения с центрипетальной ксилемой. Флоэмная часть каждого сосудистого пучка, составляющего проводящую систему тычиночной нити, развита относительно слабее ксилемы, хотя в общем проводящий пучок тычиночной нити довольно значителен. В плоских тычиночных нитях сосудистая система представлена несколькими пучками. Лубяных волокон и прочих механических элементов в тычиночных нитях обычно не встречается.
Вообще структура тычиночной нити весьма примитивна, в ней нет специальных опорных тканей и в прямом состоянии тычинка держится благодаря тургору клеток.
Значительно сложнее построен пыльник.
Развитие и строение пыльника и пыльцы будет более подробно рассмотрено в следующем параграфе. Совокупность тычинок в цветке, обычно расположенных в два круга, или цикла (мутовки), в морфологии называют андроцеем.
Аппарат, специально предназначенный для образования мегаспор и оплодотворения, составляющий в цветке последнюю верхнюю мутовку, называется гинецеем.
Общераспространенное наименование гинецея — пестик, однако оно не всеобъемлюще и не всегда может быть применено правильно. Типичный, полностью сформированный пестик состоит из трех частей: завязи, столбика и рыльца.
Ткань молодой (до оплодотворения) завязи очень проста и вся построена почти одинаково из тонкостенной паренхимы. На рисунке изображен разрез завязи сложноцветного — крестовника с содержащейся в ней семяпочкой, или мегаспорангием.
Между клетками боковых стенок завязи рассеяны небольшие межклетники. Толщу стенки завязи пронизывают тяжи прокамбия. Внутренний слой клеток стенки завязи ограничивает полость завязи, где формируется семяпочка. В некоторых плодах ковнутри от внутреннего эпидермиса расположен слой клеток, дающий впоследствии начало так называемому твердому, или кожистому, слою плода, например, у гороха — пергаментному слою, делающему плоды некоторых сортов жесткими.
Особенно мощный твердый слой развивается в таких плодах, как костянка и орех.
Наружный эпидермис завязи подстилает слой округлых ровных клеток, в зрелом плоде иногда превращающихся в толстостенные анатомические элементы — в клетки гиподермы.
Клетки паренхимы стенки завязи довольно долго сохраняют полумеристематическое состояние; окончательное оформление и дифференциация их начинается лишь спустя довольно значительный период времени после оплодотворения, вместе с началом созревания семян.
После оплодотворения завязь сильно разрастается, увеличивая свои первоначальные размеры в несколько раз.
Как в клетках паренхимы стенки завязи, так и в клетках наружного и внутреннего эпидермисов есть в относительно небольшом количестве хлоропласты, численность которых заметно увеличивается после оплодотворения, в особенности на первых стадиях созревания плода. Затем, при дальнейшем развитии процесса созревания количество хлоропластов в стенках завязи начинает опять уменьшаться и в зрелых плодах они нередко полностью разрушаются.
В стенках молодой завязи механические элементы или полностью отсутствуют, или развиты очень слабо.
Но после оплодотворения они начинают постепенно выявляться во всем своем разнообразии: входят в состав твердого луба сосудистых пучков, твердого слоя внутренней части стенки плода, а также выявляются в виде различного рода опорных и каменистых клеток мякоти плода.
Как на наружном, так и на внутреннем эпидермисах завязи могут образоваться устьица различной степени развитости, от вполне развитых до сильно редуцированных. Число устьиц на наружной стороне почти всегда бывает больше, чем на внутренней.
Наружный эпидермис завязи слегка кутинизирован. Внутренний эпидермис кутинизирован значительно слабее наружного и часто не имеет кутикулы. При развитии плода кутинизация эпидермисов обеих сторон резко усиливается, особенно наружного эпидермиса, где нередко формируется даже восковой слой.
Крупные сосудистые пучки в завязи тянутся по краям плодолистиков или по швам — местам их срастания, а также по спинной стороне каждого плодолистика.
Число слоев паренхимных клеток в зонах спинного пучка и брюшного шва значительно больше, чем в боковых стенках завязи, причем клетки ткани спинного пучка и брюшного шва значительно дольше сохраняют свойство меристематичности.
На рисунке изображен фрагмент поперечного разреза завязи в области брюшного шва одного из культурных сортов гороха. Ткань этой части завязи состоит из округлых клеток, плотно сомкнутых между собой, без значительных межклетников, т. е. несет еще черты меристематичности, причем в швах уже начинают дифференцироваться первые сосуды.
Как почти все сосуды, возникающие в первую очередь, первые сосуды швов узкие, со спиральными утолщениями и вполне соответствуют первичной ксилеме осевых органов.
Сосуды либо рассеяны поодиночке, либо собраны группами по 3—5. Механических элементов в завязи нет даже в области швов.
От брюшного шва отходят плацентарные выросты, на которых сидят ножки семяпочек, или фуникулусы. Структура спинного пучка во многом сходна со структурой брюшного: также долго сохраняется меристематичность клеток, и таким же порядком начинают дифференцироваться первые сосуды.
На протяжении развития завязи резко различаются два периода, границу между которыми составляет оплодотворение.
До оплодотворения в завязи более или менее оформлены лишь эпидермис, паренхима стенки завязи и несколько мелких групп сосудистых элементов примитивной структуры в области шва и спинного пучка. Весь несложный комплекс тканей завязи имеет свойства меристематичности. Так продолжается до оплодотворения. Если оплодотворения не произойдет или вообще семена не начнут развиваться, ткани завязи, лишенные соответствующего стимула, в большинстве случаев отмирают, завязь засыхает и отпадает. Если же оплодотворение произойдет и семена начнут развиваться, в тканях завязи начинаются энергичные процессы разрастания и дифференциации — завязь вместе с семенами превращается в плод.
Ткань столбика несколько сложнее ткани завязи, к которой столбик прикреплен.
Паренхимная ткань столбика состоит из вполне оформившихся округлых клеток с развитой системой мелких межклетников. Клетки механической ткани толстостенные, одревесневшие.
Эпидермальные клетки плотно сомкнуты друг с другом, у них заметно утолщена наружная стенка и хорошо развита кутикула.
Сосудистые пучки столбика включают твердый луб, входящий в состав полосы из механических элементов, примыкающей к эпидермису спинной стороны столбика.
Со стороны ксилемы сосудистых пучков по длине столбика тянется особый тяж, состоящий из продуктов ослизнения паренхимных клеток. Этот тяж по существу представляет как бы лизигенно образовавшееся вместилище или канал, открывающийся с одной стороны в ткани рыльца, а с другой — в полость завязи.
Если гинецей составлен несколькими плодолистиками (гинецей синкарпного плода), то канал образуется путем ослизнения полос внутреннего эпидермиса каждого из обращенных друг к другу плодолистиков. Таких заполненных слизью каналов может быть один или больше в зависимости от строения завязи и самого столбика.
Канал служит путем продвижения пыльцевой трубки к семяпочкам. Слизистое вещество, заполняющее канал, идет на питание пыльцевой трубки при ее продвижении по столбику. У многих видов канал столбика выполнен особой проводниковой тканью.
Столбик оканчивается рыльцем. Рыльца бывают различной величины и формы. Особенно своеобразно устроена поверхность рылец: эпидермис рылец образует выросты различной длины и формы, называемые сосочками и трихомами.
У одних растений сосочки очень нежные, железистые; ко времени попадания на них пыльцы они выделяют особый липкий экскрет, способствующий прилипанию к рыльцу пыльцы. В этом экскрете содержится, по-видимому, еще какое-то вещество, возможно ферментативного характера, благоприятствующее прорастанию пыльцы. Комплекс сосочков рыльца представляет собой железистую ткань.
У других растений, как, например, у гороха, вместо сосочков на рыльцах образуются настоящие волоски, причем волоски двух типов — длинные и короткие.
Оболочки длинных и коротких волосков нередко одревесневают, волоски покрывают рыльце в виде своеобразной жесткой щеточки, и среди них легко задерживается пыльца.
Нередко рыльца покрыты железистой тканью, выделяющей липкую жидкость для удержания пыльцевых зерен без сосочков и волосков.
В природе множество переходов между тремя намеченными типами поверхности рылец.
Иногда на поверхности рыльца открывается один или несколько каналов, идущих от завязи через столбик.
При отсутствии каналов пыльцевые трубки продвигаются по межклетникам и межклетным ходам. У многих растений столбик и рыльца эфемерны, после оплодотворения отмирают и высыхают.
У некоторых растений столбик и рыльце настолько прочны, что остаются на завязи до созревания плода, опадая вместе с ним. В ряде случаев столбик даже разрастается, одревесневает, превращаясь в колючку или прицепку при плоде.
В общем весь цикл своего развития столбик быстрым темпом проходит до оплодотворения.
У завязи же, наоборот, наибольшее разнообразие структуры начинает выявляться лишь после оплодотворения, при развитии семян.
Существенный объем тома занимают статьи о растениях и животных, в которых приводятся сведения об их систематическом положении, местах обитания, биологических и других особенностях.
Статьи интересны и оригинальны, т.к. в большинстве случаев сохранен авторский стиль изложения материала. Книга содержит обширную информацию по общей биологии, ботаники, зоологии, анатомии и физиологии растений, животных и человека, антропологии, палеонтологии, экологии и др. Большое внимание уделяется научным проблемам биологии.
мужской генеративный орган цветка. Состоит из тычиночной нити и пыльника. В цветке располагаются по спирали или мутовками вокруг пестика.
Тычинка, состоящая из одного пыльника, называется сидячей. Пыльник обычно состоит из двух половинок, соединённых связником. В каждой половине находятся две полости – пыльцевые гнёзда или мешки с пыльцой.
При созревании пыльцы пыльники вскрываются продольной щелью.
Иногда пыльца высыпается через дырочки или клапаны на верхушке пыльника. Самые длинные пыльники – у ветроопыляемых растений.
У некоторых растений (напр., у льна) часть тычинок не имеет пыльников. В семействе мальвовых нити тычинок срастаются, образуя характерную трубку. Ил. при ст. Цветок.
Формирование тычинки и пыльника — единый процесс развития сложной интегрированной системы. На ранних стадиях развития цветка закладывается околоцветник. Первой формируется наружная цветковая чешуя, затем — внутренняя, и почти одновременно с последней закладываются тычинки в нижних цветках колосков (рис. 4, А, Б). В верхних цветках колоска заложение тычиночных бугорков начинается позднее (рис. 4, В), когда в тычинках нижних цветков происходит мейоз. Развитие тычинки у Triticum aestivum L. — от заложения ее до формирования зрелой пыльцы — протекает в среднем в течение 3 нед.
В развитии пыльника различают 3 периода: премейотический, мейотический и постмейотический. В первый период пыльник характеризуется митотической активностью; формируются стенка пыльника и пыльцевые гнезда. В мейотический период происходят дальнейшая дифференциация слоев стенки пыльника и мейоз в микроспороцитах (материнских клетках микроспор), в результате чего образуются тетрады гаплоидных микроспор. В постмейотический период созревает пыльца, и, как правило, идет дегенерация отдельных слоев стенки пыльника.
Различают сформированную стенку гнезда пыльника, характеризующуюся окончательным, определенным числом слоев, специфичных для каждого таксона, и стенку гнезда зрелого пыльника — в момент вскрывания.
При созревании пыльника идет процесс дифференциации и специализации клеток всех слоев его стенки, в основном совпадающий с мейотическим периодом.
На первых этапах развития тычиночный бугорок состоит из меристематических клеток. Одновременно с обособлением проводящего пучка на поперечном срезе пыльника можно наблюдать образование 4 его лопастей (гнезд). Каждая лопасть преобразуется в микроспорангий. 2 латеральных гнезда составляют теку. Теки соединены между собой связником.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
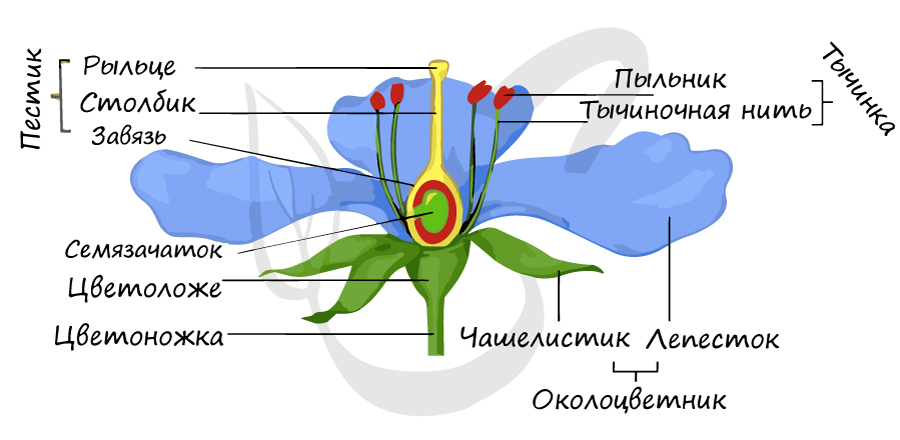
Цветок представляет собой укороченный видоизмененный побег покрытосеменных растений, специализированный для образования спор и гамет, а также для осуществления полового процесса, результатом которого является развитие плода с семенами.
Строение цветка
Приступим к классификации частей цветка. Цветок состоит из:
- Стеблевой части, в которой выделяется:
- Цветоножка - разветвление стебля, на котором расположен цветок
- Цветоложе - расширенная верхняя часть цветоножки, от которой отходят чашелистики, лепестки, тычинки, пестики
- Чашелистики - видоизмененные листья, составляющие чашечку листа
- Лепестки - внутренние видоизмененные листья, составляющие венчик листа
Отметим, что в ботанике есть такое понятие как околоцветник: так называют внешнюю часть цветка, окружающую репродуктивные органы. Обычно околоцветник состоит из внешнего кольца чашелистиков (чашечка) и внутреннего кольца лепестков (венчик).
- Тычинки - мужской половой орган цветка, состоящий из тычиночной нити и пыльника, в гнездах которого образуется пыльца. Каждое пыльцевое зерно содержит 2 гаплоидные клетки: вегетативную и генеративную.
- Пестик - основная расположенная в центре часть цветка, является женским половым органом.
Состоит из завязи - нижней утолщенной части пестика, из которой в дальнейшем образуется плод, столбика - центральной части пестика между завязью и рыльцем, и самого рыльца - широкой верхней части пестика, на которую попадает пыльца.
В завязи пестика формируются семязачатки, которые после опыления и оплодотворения образуют семена. Выделяют цветки с верхней завязью - картофель, горох, редька, гвоздика и с нижней завязью - у огурцов, колокольчиков, подсолнечника. Верхняя завязь свободная, ее легко выделить из цветка. Выделить нижнюю завязь, не повредив цветок, значительно труднее, так как она срастается с тычинками, листами околоцветника и даже с цветоложем (у огурца).
![Строение цветка]()
Особо отметьте наличие в цветке нектарников (медовиков). Они привлекают насекомых-опылителей, выделяя нектар - сахаристый сок с характерным запахом. При попытке собрать нектар насекомые сотрясают генеративную часть цветка, рассыпая пыльцу на себя, на рыльце пестика (благодаря чему происходит опыление) и на другие части цветка. Сами насекомые служат опылителями, перенося на тельце и конечностях пыльцу с одних цветков на другие.
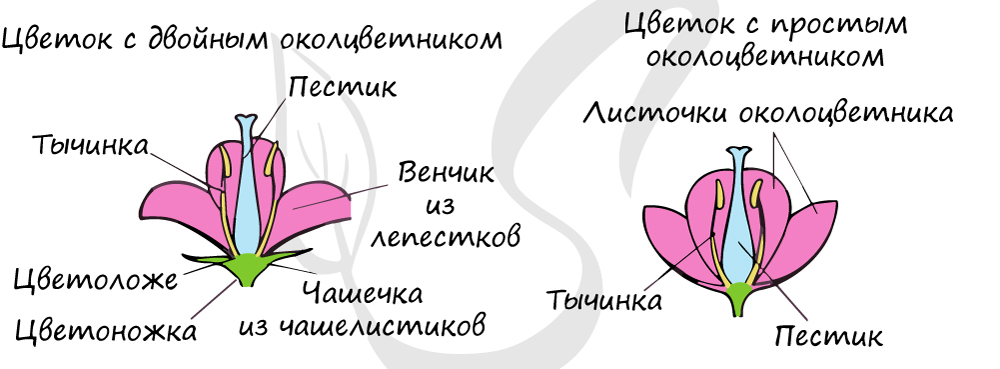
Околоцветник
Вместе чашелистики и лепестки составляют околоцветник. Околоцветник цветка бывает двойным и простым. Двойной околоцветник включает в себя чашечку и венчик, имеется у яблони, гороха, картофеля. Если околоцветник не разделен на чашечку и венчик, то его называют простым. Простой околоцветник состоит из листочков, характерен для лука, дуба, березы, тюльпана и ландыша. У некоторых растений околоцветник отсутствует, их цветки называются "голые" : у тополя, вербы.
![Двойной и простой околоцветник]()
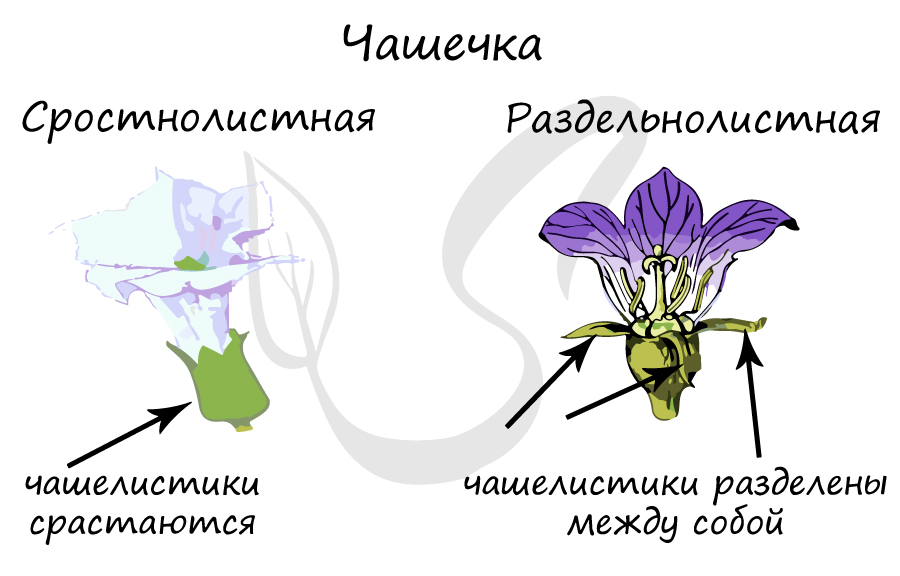
Чашечка
- Раздельнолистную чашечку - состоит из разделенных между собой чашелистиков: у дикой редьки, земляники
- Сростнолистная чашечка - чашелистики сращены между собой: у гвоздики, гороха
![Раздельнолистная и сростнолистная чашечка]()
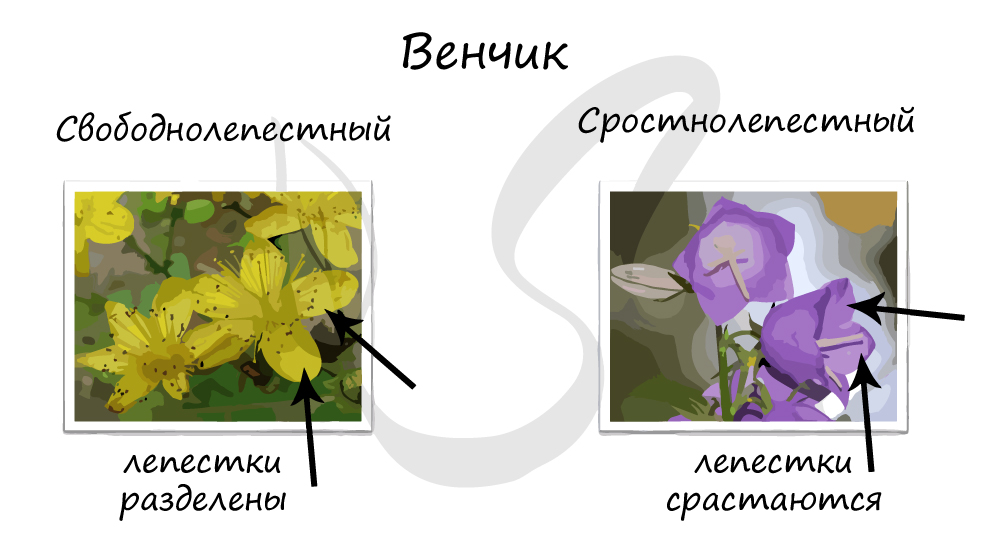
Венчик
- Свободнолепестный - лепестки венчика разделены между собой
- Спайнолепестный - лепестки венчика срастаются друг с другом
В дальнейшем по мере изучения семейств покрытосеменных мы изучим формулы цветков. Запомните сейчас, что в случае, если любые части цветка срастаются между собой, то в формуле цветка их число берется в скобки.
![Свободнолепестный и спайнолепестный венчик]()
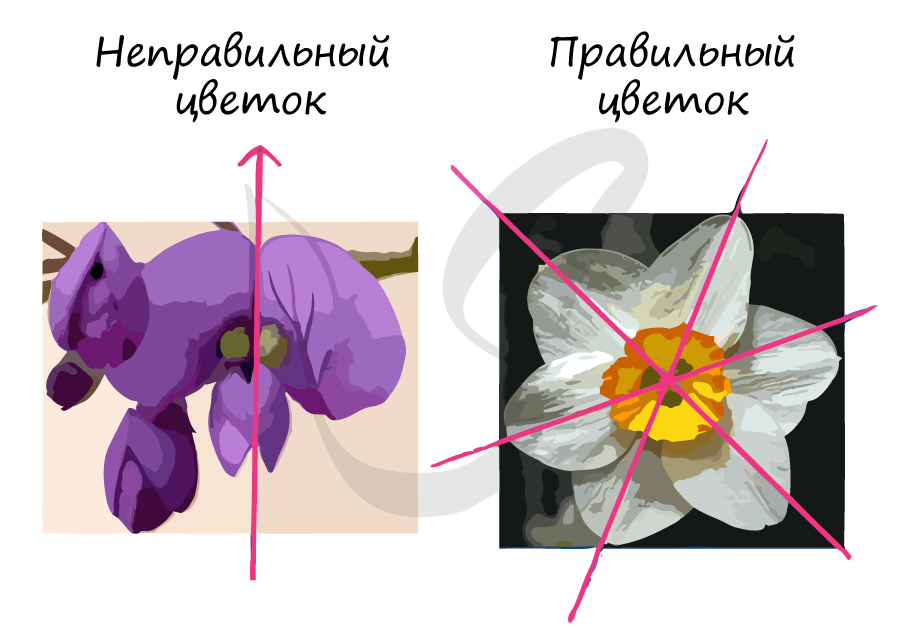
Симметрия цветка
- Правильные (актиноморфные), через которые можно провести множество плоскостей симметрии. Правильные цветки имеются у гвоздики, лилии, огурцов. В формуле такие цветки обозначаются знаком *
- Неправильные (зигоморфные), такие цветки имеют только одну плоскость симметрии. Цветки такого типа есть у гороха, шалфея, львиного зева. В формуле такой цветок обозначается знаком ↑
![Правильные и неправильные цветки у растений]()
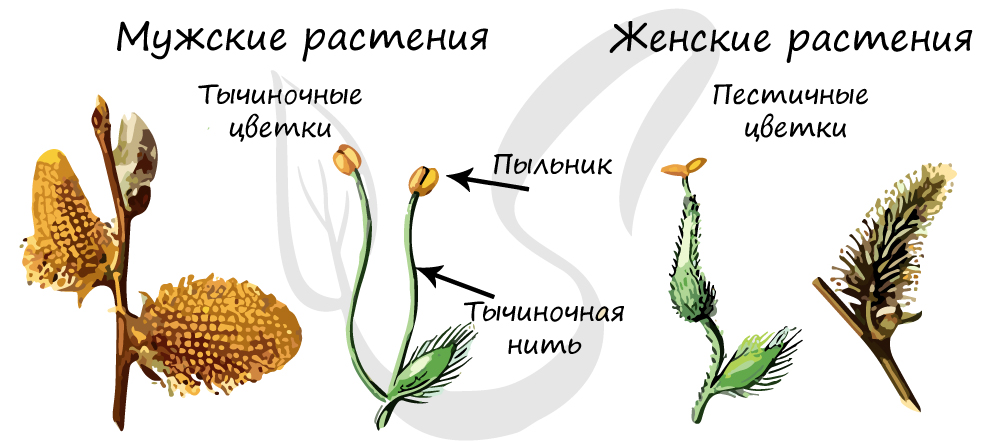
Однодомные и двудомные растения
Обоеполые цветки имеют и тычинки, и пестики в одном цветке. Однако есть растения, у которых тычинки и пестики расположены на разных цветках. У таких растений на цветке находятся либо тычинки (тычиночные цветки) - мужские цветки, либо пестики (пестичные) - женские цветки. В зависимости от расположения мужских и женских цветков эти растения делятся на:
- Однодомные - у них и мужские, и женские цветки расположены на одном и том же растении: у кукурузы, березы, тыквы.
- Двудомные - имеют и женские, и мужские цветки, расположенные на разных растениях: у тополя, конопли, вербы.
Поделюсь своей собственной ассоциацией, чтобы вы успешно запомнили эти понятия. Вообразите, что в гости к зажиточным хозяевам приехало большое количество гостей. Богатые хозяева построили на участке два дома, и у них есть возможность разделить всех гостей, так что мужчины отделяются от женщин и идут в разные дома ("двудомные растения"). В случае если хозяева оказались менее богаты, то у них только один дом, так что гостям и мужского, и женского пола придется искать место для ночевки в одном доме ("однодомные растения").
![Однодомные и двудомные растения]()
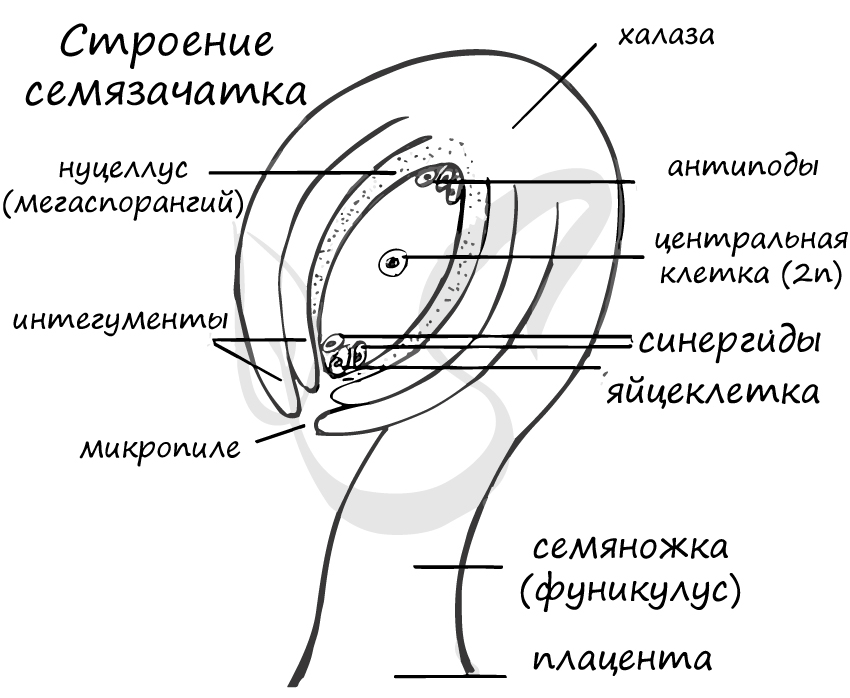
Семязачаток
Также называется семяпочкой. Представляет собой образующийся в завязи многоклеточный орган, из которого развивается семя. Ткани завязи образуют выступ (вырост), называющийся плацента, которым семязачаток крепится внутри завязи. С помощью семяножки семязачаток сообщается с плацентой.
![Строение семязачатка (семяпочки)]()
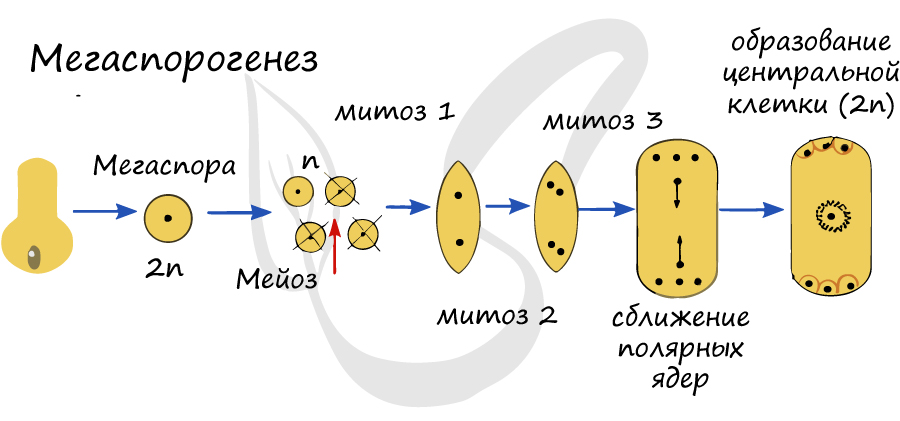
Процесс локализуется в нуцеллусе, называющимся мегаспорангием. Материнская клетка (2n) начинает делиться мейозом, и, что предсказуемо, получается четыре клетки - четыре гаплоидные мегаспоры (n). Из них три отмирают, выживает только одна, приближенная к халазе - ткани, где соединяются интегумент и нуцеллус.
Запомните, что из мегаспоры развивается женский гаметофит - зародышевый мешок. Гаметофит у растений это гаплоидная многоклеточная фаза в цикле развития, которая чередуется со спорофитом - диплоидной фазой.
Ядро мегаспоры трижды делится эндомитозом (удвоение числа хромосом внутри ядерной оболочки, без разрушения ядрышка и без образования нитей веретена деления). В результате образуется 8 ядер, по 4 ядра у каждого полюса зародышевого мешка. На этой восьмиядерной стадии деление ядра женского гаметофита окончено.
От каждого из двух полюсов в центр зародышевого мешка направляется по одному ядру, так называемые - полярные ядра. Таким образом, у полюсов зародышевого мешка их остается по три. Две клетки в центре сливаются и образуют центральную клетку, диплоидного (2n) набора хромосом. На микропилярном полюсе зародышевого мешка одна наиболее крупная клетка превращается в яйцеклетку, а две других становятся вспомогательными клетками - синергидами, короткоживущими клетками. Вместе яйцеклетка и синергиды образуют яйцевой аппарат.
![Мегаспорогенез]()
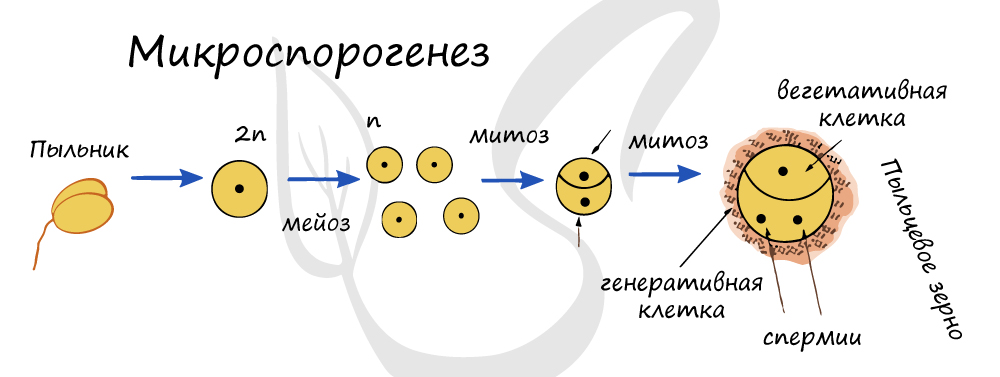
Локализуется в микроспорангиях - гнездах пыльника. Диплоидная материнская клетка делится мейозом, в результате образуется четыре микроспоры с гаплоидным набором хромосом. Каждая из микроспор делится митозом, в результате получаются две клетки: крупная вегетативная и более мелкая генеративная - эти две клетки и составляют пыльцевое зерно (пыльцу). Пыльцевое зерно состоит из двух оболочек - интины (внутренней) и экзины (наружной).
Важно отметить, что из генеративной клетки к моменту оплодотворения (еще в пыльнике (до опыления) или в пыльцевой трубке (после опыления)) путем митоза образуются мужские половые клетки - спермии (или сперматозоиды), необходимые для процесса оплодотворения. Запомните, мужской гаметофит семенного растения - пыльцевое зерно.
![Микроспорогенез]()
Опыление
Признаки самоопыляющихся растений: запах и нектар отсутствуют, тычинки выше пестиков, иногда пыльца созревает еще в бутоне и опыление происходит в цветке еще до его распускания.
Перенос пыльцы из пыльника цветка одного растения на рыльце пестика другого растения. Отметим искусственное опыление, которое сознательно осуществляет человек для повышения урожайности или выведения новых сортов. Осуществляется с помощью воды, ветра и животных. Здесь необходимо ввести новые термины:
Такие растения имеют следующие характерные черты: у них мелкие цветки, невзрачный околоцветник, цветки лишены нектарников (то есть запах, нектар у цветов отсутствует). Ветроопыляемые растения обычно растут большими скоплениями (заросли тростника, березовые рощи), зацветают ранней весной, до появления листьев. Тычинки располагаются на длинных, свисающих тычиночных нитях. Пыльцы образуется очень много, она мелкая, легкая и сухая.
![Ветроопыляемые растения]()
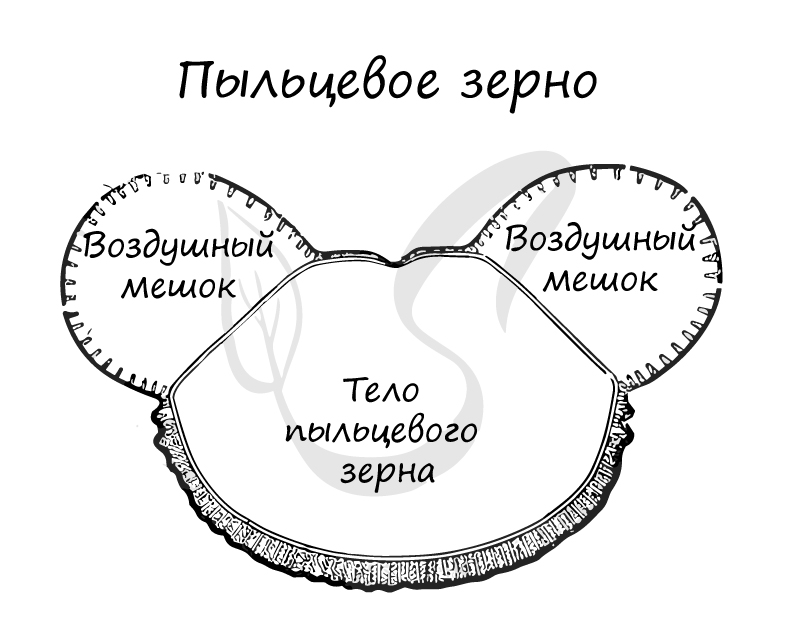
Пыльцевые зерна благодаря наличию воздушных мешков могут перемещаться на большие расстояния, достигающие десятков километров: 30-35 км у березы, у ольхи до 400 км.
![Воздушные мешки пыльцевого зерна]()
Эти растения отличают крупные цветки, мелкие - собраны в соцветия. Имеют нектарники и характерный запах (аромат), особенно важный для привлечения насекомых. Пыльцы мало, она крупная, тяжелая, липкая. Ее внешний слой (экзина) часто покрыт различными приспособлениями, которые помогают зацепится за насекомых: бугорки, шипы, гребешки.
Теперь вы точно знаете, почему именно насекомооплыяемые растения стоит дарить прекрасным девушкам, а не ветроопыляемые (на первом свидании точно лучше подстраховаться насекомооплыяемыми, хотя если вы хотите удивить - вперед в березовую рощу ;)
![Насекомоопыляемые растения]()
Оплодотворение
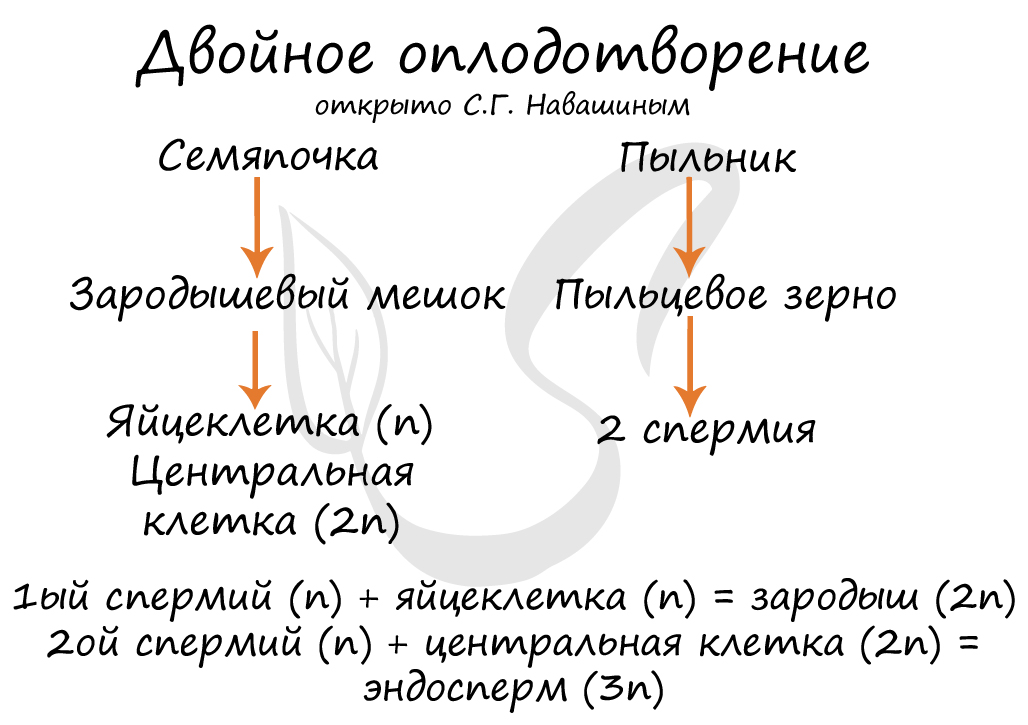
Оплодотворение - слияние спермия, сперматозоида (мужской половой клетки) с яйцом, яйцеклеткой (женской половой клеткой), приводящее к образованию зиготы. Тем или иным способом пыльца (пыльцевое зерно) оказывается на рыльце пестика. Вегетативная клетка начинает прорастать в ткани пестика, растворяя их, формирует пыльцевую трубку. Из генеративной клетки образуются два спермия.
Пыльцевая трубка прорастает до зародышевого мешка, благодаря чему спермии достигают яйцеклетки. Далее у цветковых растений происходит уникальное явление, открытое С.Г. Навашиным - двойное оплодотворение. Как вы помните, из генеративной клетки образовалось два спермия. Суть двойного оплодотворения заключается в том, что один из спермиев сливается с яйцеклеткой (оплодотворяет ее) с образованием зиготы (диплоидна), из которой развивается зародыш. Второй спермий сливается с центральной клеткой (эта клетка к моменту слияния уже диплоидна) с образованием эндосперма (триплоиден) - запасного питательного вещества.
![Насекомоопыляемые растения]()
После оплодотворения с течением времени из семязачатков образуются семена. Из интегумента семязачатка (от лат. integumentum — покрывало, покров) образуется семенная кожура. Околоплодник формируется из стенок завязи пестика.
Соцветия
Цветки, особенно у насекомооплыемых растений, редко расположены по одиночке. Чаще всего цветки образуют скопления - соцветия. Соцветие - часть годичного побега растения, несущая цветки и видоизмененные прицветные листья, в пазухах которых и располагаются цветки или соцветия.
Простыми называют соцветия с одной осью - главной, на которой расположены цветки. К простым соцветиям относятся:
-
Кисть - цветки поочередно крепятся к неразветвленной удлиненной главной оси. Имеется у ландыша, черемухи.
![Соцветие кисть ландыша]()
![Соцветие щиток груши]()
![Соцветие колос подорожника]()
![Соцветие початок кукурузы]()
![Соцветие корзинка одуванчика]()
![Соцветие зонтик примулы]()
![Соцветие головка клевера]()
Сложными называют соцветия, у которых на главной оси расположены не цветки, а частные (парциальные) соцветия.
-
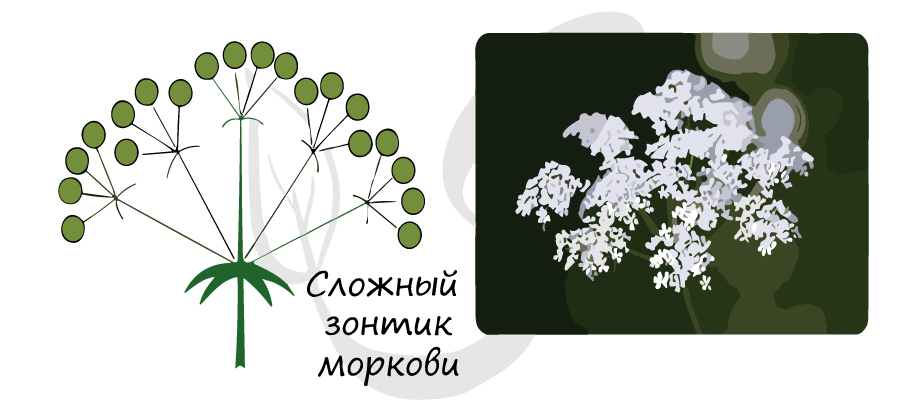
Метелка - по-другому называется - сложная кисть. Главная ось ветвится, от нее отходят оси боковые, на которых расположены цветки - у сирени, или колоски: у овса, риса, просо.
![Метелка соцветие]()
![Сложный зонтик моркови]()
![Сложный колос пшеницы]()
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Морфологически и функционально тычинка соответствует микроспорофиллу. Существенной частью тычинки является микроспорангий — пыльник, в котором образуются микроспоры — пыльца.
Тычинка состоит из тычиночной нити, связника и пыльников. Обычно образуется два пыльника, в каждом из которых имеется два пыльцевых гнезда. Однако в некоторых случаях пыльцевых гнезд может быть меньше или больше. Связник — осевая часть тычинки, которая как бы является продолжением тычиночной нити и соединяет между собой пыльники.
Хорошо развитые тычинки не обнаруживают внешнего сходства с листом. Но это сходство проявляется в некоторых чертах анатомического строения и в процессе онтогенеза. Некоторые примитивные тычинки имеют расширенную пластинчатую нить, отдаленно напоминающую лист. В этом случае увеличивается и анатомическое сходство с листом.
Тычинки обнаруживают большое разнообразие формы, что обусловлено способом прикрепления пыльников и характером их раскрывания.
Строение тычиночной нити очень простое. Снаружи она покрыта эпидермисом, клетки которого слегка вытянуты, имеют кутикулу. Устьица на тычиночной нити обычно отсутствуют. Под эпидермисом располагается мезофилл, состоящий из немногих округлых тонкостенных клеток. Центральное положение в тычиночной нити занимает проводящий пучок.
Пыльник имеет многослойные стенки, ограничивающие пыльцевое гнездо. Наружным слоем служит эпидермис, состоящий из мелких плоских клеток с хорошо развитой кутикулой. Под эпидермисом располагается фиброзный слой. Клетки его характеризуются неравномерным утолщением стенок. Размеры, форма клеток этого слоя, характер утолщений в клеточных оболочках различны у разных растений. Фиброзный слой способствует раскрыванию пыльника. У некоторых растений он отсутствует.
Самый внутренний слой в стенках пыльника — это выстилающий слой, или тапетум. Он непосредственно выстилает пыльцевое гнездо и участвует в питании созревающей пыльцы. Клетки тапетума относительно крупные, тонкостенные, богатые зернистой цитоплазмой. Они часто бывают многоядерными или содержат полиплоидные ядра. У большинства растений тапетум — недолговечная ткань. Ко времени созревания спор клетки тапетума разрушаются. Цитоплазма их сливается, образуя тапетальный плазмодий, который распространяется в пыльцевом гнезде. Продукты растворения тапетума содержат много Сахаров и жиров, смешанных со слизистыми веществами.
Пыльцевое гнездо заполнено в зависимости от степени развития пыльника пыльцой или материнскими клетками пыльцы, или тканью, предшествующей развитию материнских клеток, — археспорием.
Связник состоит из паренхимной ткани и имеет проводящий пучок, который является продолжением проводящего пучка тычиночной нити. Иногда проводящий пучок в связнике ветвится. У разных растений связник развит в разной степени.
Развитие тычинки начинается с образования на конусе нарастания небольшого бугорка. В центре его обособляются крупные, богатые цитоплазмой клетки — это материнские, или первичные, клетки археспория. В результате деления первичных клеток археспория тангентальными перегородками обособляется тапетум, образующийся из наружного слоя клеток. Из внутреннего слоя возникают вторичные клетки археспория, или материнские клетки пыльцы.
Пыльца образуется из вторичных клеток археспория путем их редукционного деления (мейоза). Каждая материнская клетка дает четыре гаплоидные — тетраду спор. Это и есть клетки пыльцы, или пыльцевые зерна. Каждое пыльцевое зерно представляет собой гаплоидную клетку с двухслойной оболочкой. Наружный слой оболочки называется экзиной, внутренний — интиной. Экзина более плотная, имеет часто местные утолщения, которые придают пыльцевой поверхности характерный вид. В экзине имеются поры. Интина, более тонкая и мягкая часть оболочки, состоит из пектина.
Тычи́нка (лат. Stamen, устар. пыльцелистик[1]) — часть цветкацветковых растений, мужской репродуктивный орган, в котором образуется пыльца.
Строение тычинки
Тычинка состоит из тычиночной нити (стерильной части) и пыльника, в котором образуются пыльцевые зёрна. Тычиночная нить является гомологоммикроспорофилла разноспоровых плауновидных и голосеменных[2].
В каждом из четырёх гнёзд пыльника из клеток археспория после мейоза образуются микроспоры, которые затем прорастают в мужские гаметофиты — пыльцевые зёрна. Совокупность пыльцевых зёрен называется пыльцой[2].
Число тычинок в одном цветке может быть от одной до нескольких тысяч (например, у некоторых кактусов). Совокупность тычинок цветка называется андроцеем. Тычинки обычно расположены на цветоложе спирально или по кругу.
Тычинки могут полностью или частично срастаться.
У тыквенных они срастаются полностью, у астровых (сложноцветных) — пыльниками, у бобовых — тычиночными нитями (сросшиеся тычиночные нити в цветках представителей бобовых называются тычиночными трубками[2]).
У раффлезиевых и ряда представителей орхидных тычинки срастаются с пестиком, образуя особый орган, называемый колонкой.
Каждая тычинка в типичном случае состоит из двух основных частей: стерильной — тычиночной нити, или филаментума, и фертильной — пыльника, или микроспорангия.
Тычиночная нить у некоторых растений может отсутствовать и пыльники таких тычинок называются сидячими.
У некоторых растений тычиночные нити срастаются между собой, как, например, у зверобойных, бобовых, у других — ветвятся, как у клещевины.
Эпидермис тычиночной нити состоит из удлиненных по оси клеток с тонкой кутикулой, а нередко и совсем без кутикулы. Клетки эпидермиса тычиночной нити соединены между собой не столь плотно, как клетки эпидермиса листьев.
Устьица на тычиночных нитях встречаются иногда в значительном числе. Мезофилл нормальной тычиночной нити состоит из паренхимных клеток с тонкими оболочками и хорошо выраженными межклетниками. Многочисленные и обширные межклетники и тонкость оболочек клеток придают мягкость и рыхлость структуре-мезофилла тычиночных нитей.
В центре типичной тычиночной нити расположена сосудистая система первичного строения с центрипетальной ксилемой. Флоэмная часть каждого сосудистого пучка, составляющего проводящую систему тычиночной нити, развита относительно слабее ксилемы, хотя в общем проводящий пучок тычиночной нити довольно значителен. В плоских тычиночных нитях сосудистая система представлена несколькими пучками. Лубяных волокон и прочих механических элементов в тычиночных нитях обычно не встречается.
Вообще структура тычиночной нити весьма примитивна, в ней нет специальных опорных тканей и в прямом состоянии тычинка держится благодаря тургору клеток.
Значительно сложнее построен пыльник.
Развитие и строение пыльника и пыльцы будет более подробно рассмотрено в следующем параграфе. Совокупность тычинок в цветке, обычно расположенных в два круга, или цикла (мутовки), в морфологии называют андроцеем.
Аппарат, специально предназначенный для образования мегаспор и оплодотворения, составляющий в цветке последнюю верхнюю мутовку, называется гинецеем.
Общераспространенное наименование гинецея — пестик, однако оно не всеобъемлюще и не всегда может быть применено правильно. Типичный, полностью сформированный пестик состоит из трех частей: завязи, столбика и рыльца.
Ткань молодой (до оплодотворения) завязи очень проста и вся построена почти одинаково из тонкостенной паренхимы. На рисунке изображен разрез завязи сложноцветного — крестовника с содержащейся в ней семяпочкой, или мегаспорангием.
Между клетками боковых стенок завязи рассеяны небольшие межклетники. Толщу стенки завязи пронизывают тяжи прокамбия. Внутренний слой клеток стенки завязи ограничивает полость завязи, где формируется семяпочка. В некоторых плодах ковнутри от внутреннего эпидермиса расположен слой клеток, дающий впоследствии начало так называемому твердому, или кожистому, слою плода, например, у гороха — пергаментному слою, делающему плоды некоторых сортов жесткими.
Особенно мощный твердый слой развивается в таких плодах, как костянка и орех.
Наружный эпидермис завязи подстилает слой округлых ровных клеток, в зрелом плоде иногда превращающихся в толстостенные анатомические элементы — в клетки гиподермы.
Клетки паренхимы стенки завязи довольно долго сохраняют полумеристематическое состояние; окончательное оформление и дифференциация их начинается лишь спустя довольно значительный период времени после оплодотворения, вместе с началом созревания семян.
После оплодотворения завязь сильно разрастается, увеличивая свои первоначальные размеры в несколько раз.
Как в клетках паренхимы стенки завязи, так и в клетках наружного и внутреннего эпидермисов есть в относительно небольшом количестве хлоропласты, численность которых заметно увеличивается после оплодотворения, в особенности на первых стадиях созревания плода. Затем, при дальнейшем развитии процесса созревания количество хлоропластов в стенках завязи начинает опять уменьшаться и в зрелых плодах они нередко полностью разрушаются.
В стенках молодой завязи механические элементы или полностью отсутствуют, или развиты очень слабо.
Но после оплодотворения они начинают постепенно выявляться во всем своем разнообразии: входят в состав твердого луба сосудистых пучков, твердого слоя внутренней части стенки плода, а также выявляются в виде различного рода опорных и каменистых клеток мякоти плода.
Как на наружном, так и на внутреннем эпидермисах завязи могут образоваться устьица различной степени развитости, от вполне развитых до сильно редуцированных. Число устьиц на наружной стороне почти всегда бывает больше, чем на внутренней.
Наружный эпидермис завязи слегка кутинизирован. Внутренний эпидермис кутинизирован значительно слабее наружного и часто не имеет кутикулы. При развитии плода кутинизация эпидермисов обеих сторон резко усиливается, особенно наружного эпидермиса, где нередко формируется даже восковой слой.
Крупные сосудистые пучки в завязи тянутся по краям плодолистиков или по швам — местам их срастания, а также по спинной стороне каждого плодолистика.
Число слоев паренхимных клеток в зонах спинного пучка и брюшного шва значительно больше, чем в боковых стенках завязи, причем клетки ткани спинного пучка и брюшного шва значительно дольше сохраняют свойство меристематичности.
На рисунке изображен фрагмент поперечного разреза завязи в области брюшного шва одного из культурных сортов гороха. Ткань этой части завязи состоит из округлых клеток, плотно сомкнутых между собой, без значительных межклетников, т. е. несет еще черты меристематичности, причем в швах уже начинают дифференцироваться первые сосуды.
Как почти все сосуды, возникающие в первую очередь, первые сосуды швов узкие, со спиральными утолщениями и вполне соответствуют первичной ксилеме осевых органов.
Сосуды либо рассеяны поодиночке, либо собраны группами по 3—5. Механических элементов в завязи нет даже в области швов.
От брюшного шва отходят плацентарные выросты, на которых сидят ножки семяпочек, или фуникулусы. Структура спинного пучка во многом сходна со структурой брюшного: также долго сохраняется меристематичность клеток, и таким же порядком начинают дифференцироваться первые сосуды.
На протяжении развития завязи резко различаются два периода, границу между которыми составляет оплодотворение.
До оплодотворения в завязи более или менее оформлены лишь эпидермис, паренхима стенки завязи и несколько мелких групп сосудистых элементов примитивной структуры в области шва и спинного пучка. Весь несложный комплекс тканей завязи имеет свойства меристематичности. Так продолжается до оплодотворения. Если оплодотворения не произойдет или вообще семена не начнут развиваться, ткани завязи, лишенные соответствующего стимула, в большинстве случаев отмирают, завязь засыхает и отпадает. Если же оплодотворение произойдет и семена начнут развиваться, в тканях завязи начинаются энергичные процессы разрастания и дифференциации — завязь вместе с семенами превращается в плод.
Ткань столбика несколько сложнее ткани завязи, к которой столбик прикреплен.
Паренхимная ткань столбика состоит из вполне оформившихся округлых клеток с развитой системой мелких межклетников. Клетки механической ткани толстостенные, одревесневшие.
Эпидермальные клетки плотно сомкнуты друг с другом, у них заметно утолщена наружная стенка и хорошо развита кутикула.
Сосудистые пучки столбика включают твердый луб, входящий в состав полосы из механических элементов, примыкающей к эпидермису спинной стороны столбика.
Со стороны ксилемы сосудистых пучков по длине столбика тянется особый тяж, состоящий из продуктов ослизнения паренхимных клеток. Этот тяж по существу представляет как бы лизигенно образовавшееся вместилище или канал, открывающийся с одной стороны в ткани рыльца, а с другой — в полость завязи.
Если гинецей составлен несколькими плодолистиками (гинецей синкарпного плода), то канал образуется путем ослизнения полос внутреннего эпидермиса каждого из обращенных друг к другу плодолистиков. Таких заполненных слизью каналов может быть один или больше в зависимости от строения завязи и самого столбика.
Канал служит путем продвижения пыльцевой трубки к семяпочкам. Слизистое вещество, заполняющее канал, идет на питание пыльцевой трубки при ее продвижении по столбику. У многих видов канал столбика выполнен особой проводниковой тканью.
Столбик оканчивается рыльцем. Рыльца бывают различной величины и формы. Особенно своеобразно устроена поверхность рылец: эпидермис рылец образует выросты различной длины и формы, называемые сосочками и трихомами.
У одних растений сосочки очень нежные, железистые; ко времени попадания на них пыльцы они выделяют особый липкий экскрет, способствующий прилипанию к рыльцу пыльцы. В этом экскрете содержится, по-видимому, еще какое-то вещество, возможно ферментативного характера, благоприятствующее прорастанию пыльцы. Комплекс сосочков рыльца представляет собой железистую ткань.
У других растений, как, например, у гороха, вместо сосочков на рыльцах образуются настоящие волоски, причем волоски двух типов — длинные и короткие.
Оболочки длинных и коротких волосков нередко одревесневают, волоски покрывают рыльце в виде своеобразной жесткой щеточки, и среди них легко задерживается пыльца.
Нередко рыльца покрыты железистой тканью, выделяющей липкую жидкость для удержания пыльцевых зерен без сосочков и волосков.
В природе множество переходов между тремя намеченными типами поверхности рылец.
Иногда на поверхности рыльца открывается один или несколько каналов, идущих от завязи через столбик.
При отсутствии каналов пыльцевые трубки продвигаются по межклетникам и межклетным ходам. У многих растений столбик и рыльца эфемерны, после оплодотворения отмирают и высыхают.
У некоторых растений столбик и рыльце настолько прочны, что остаются на завязи до созревания плода, опадая вместе с ним. В ряде случаев столбик даже разрастается, одревесневает, превращаясь в колючку или прицепку при плоде.
В общем весь цикл своего развития столбик быстрым темпом проходит до оплодотворения.
У завязи же, наоборот, наибольшее разнообразие структуры начинает выявляться лишь после оплодотворения, при развитии семян.
Существенный объем тома занимают статьи о растениях и животных, в которых приводятся сведения об их систематическом положении, местах обитания, биологических и других особенностях.
Статьи интересны и оригинальны, т.к. в большинстве случаев сохранен авторский стиль изложения материала. Книга содержит обширную информацию по общей биологии, ботаники, зоологии, анатомии и физиологии растений, животных и человека, антропологии, палеонтологии, экологии и др. Большое внимание уделяется научным проблемам биологии.
мужской генеративный орган цветка. Состоит из тычиночной нити и пыльника. В цветке располагаются по спирали или мутовками вокруг пестика.
Тычинка, состоящая из одного пыльника, называется сидячей. Пыльник обычно состоит из двух половинок, соединённых связником. В каждой половине находятся две полости – пыльцевые гнёзда или мешки с пыльцой.
При созревании пыльцы пыльники вскрываются продольной щелью.
Иногда пыльца высыпается через дырочки или клапаны на верхушке пыльника. Самые длинные пыльники – у ветроопыляемых растений.
У некоторых растений (напр., у льна) часть тычинок не имеет пыльников. В семействе мальвовых нити тычинок срастаются, образуя характерную трубку. Ил. при ст. Цветок.
Созревание тычинок и пестиков в одном и том же цветке у многих растений происходит неодновременно: чаще тычинки созревают раньше, а пестики позднее, как, например, у подсолнечника, некоторых груш, яблонь, земляники, апельсина, герани и многих других.
Противоположный случай, когда сперва пестик бывает готов к восприятию пыльцы, а потом уже созревают и вскрываются пыльники, встречается реже: например, у подорожника, белены, душистого колоска.
У гречихи, первоцвета, медуницы и некоторых других растений перекрестное опыление достигается путем разностолбчатости. Например, у гречихи на одних растениях все цветки имеют длинные столбики и короткие тычинки, а на других растениях, наоборот, короткие столбики и длинные тычинки.
Опыление у гречихи происходит преимущественно пчелами, которые собирают с гречихи обильный взяток. При посещении длинностолбчатых цветков к головке пчелы пристает пыльца с коротких тычинок, которая затем переносится пчелой на рыльца пестиков короткостолбчатых цветков. Брюшко пчелы при этом обсыпается пыльцой длинных тычинок.
При посещении других длинностолбчатых цветков пчела переносит пыльцу на рыльца этих цветков. Известно, что гречиха дает лучшие урожаи, когда в период ее цветения вблизи находятся ульи с пчелами.
Читайте также: