
Факторы устойчивости растений к повреждению фитофагами
Обновлено: 05.10.2024
Фитоиммунитет — невосприимчивость растений к патогенам, а также к насекомым, обеспечивается множеством механизмов: выработкой низкомолекулярных фитонцидов, обладающих антибактериальными и фунгицидными свойствами, рецепторами распознавания специфических белковых и углеводных последовательностей, характерных для многих возбудителей, и системой подавления экспрессии генов при помощи РНК в качестве противовирусной защиты.
Для защиты от потенциальных вредителей и болезней у растений имеется большой арсенал приспособлений. Хорошим барьером для некоторых бактерий и грибов являются кутикула и перидерма. Шипы и жгучие волоски надежно защищают растения от крупных животных, однако они имеются не у всех видов растений и бесполезны против мелких вредителей и патогенов. Главным же оружием у всех растений служит химическая система защиты, которая насчитывает тысячи разнообразных соединений вторичного происхождения. Именно вторичные соединения обеспечивают основную защиту растения от поедания животными и инфицирования патогенами.
К подавляющему большинству патогенов у растений имеется врожденный иммунитет. Почти непреодолимым барьером для многих патогенов, не обладающих специальными приспособлениями для активного проникновения, служат покровы и клеточная стенка растений. И только их повреждение может открыть паразиту доступ к внутренним тканям. Причиной видового иммунитета растения часто является его пищевая непригодность для патогена. Это может определяться наличием токсических соединений, отсутствием или низким содержанием в тканях растения веществ, например стеринов, необходимых для нормального развития паразита или фитофага. Растения обладают большим спектром токсических соединений, которые все время присутствуют в тканях независимо от того, подвергались они инфицированию или нет. Некоторые из них (например, сапонины) постоянно находятся в активной форме, другие же хранятся в виде неактивных предшественников в вакуоли (цианогенные гликозиды, глюкозинолаты) и активируются только при повреждении клеток. Синтез таких прототоксинов, которые было предложено называть фитоантисипинами, осуществляется цитоплазматическими ферментами, которые активируются при повреждении тканей патогеном. Именно фитоантисипины часто выступают в качестве первого химического барьера на пути проникновения патогена.
Одним из самых эффективных способов защиты растительных организмов от повреждения является реакция сверхчувствительности (СВЧ), т. е. быстрая локальная гибель инфицированных растительных клеток вместе с патогеном, что, в конечном счете, обеспечивает устойчивость всего растения. В том случае, когда несколько растительных клеток быстро распознают патоген и погибают вместе с ним, выживает целый организм. Поэтому самым важным этапом во взаимодействии растения с патогеном является процесс распознавания паразита. Специфичность взаимодействия определяется белковыми продуктами генов устойчивости (R-генов) растения-хозяина и генов авирулентности патогена. Каждому гену авирулентности патогена соответствует свой ген устойчивости растения.
Окислительный взрыв – процесс образования большого количества активных форм кислорода (АФК) и свободных радикалов на поверхности клетки. Этот процесс является одной из наиболее ранних реакций практически на любые стрессовые воздействия (как биотической, так и абиотической природы). В реакции СВЧ окислительный взрыв завершается появлением некротических пятен и гибелью части клеток растения-хозяина, а также патогена. При этом в некротических участках формируются механические барьеры, препятствующие распространению патогена по тканям.
Синтез фитогормонов. Образование при окислительном взрыве больших количеств пероксида водорода также является причиной активации синтеза фитогормонов — салициловой и жасмоновой кислот. Повышение содержания салициловой кислоты (СК) усиливает реакцию СВЧ, поскольку салицилат является ингибитором каталазы — фермента, расщепляющего пероксид водорода. То есть пероксид водорода, активируя синтез салициловой кислоты, способствует еще большему накоплению активных форм кислорода и таким образом вызывает усиление СВЧ-реакции. Жасмоновая кислота активирует экспрессию ряда генов, продукты которых позволяют растениям адаптироваться к патогенам и механическим повреждениям. К их числу относятся токсичные для патогенов низкомолекулярные цистеин-богатые белки тионины и полипептиды дефензины, ферменты, участвующие в синтезе фитоалексинов (большой группы метаболитов вторичного происхождения, которые подавляют развитие патогена).
При инфицировании растения в каком-либо одном месте очень часто происходит формирование устойчивости к последующему нападению во всех тканях и органах организма. Это явление было названо системной приобретенной устойчивостью (СПУ), или системным приобретенным иммунитетом. Важным свойством системного иммунитета является быстрота его появления. При повторном заражении некротические пятна появляются гораздо быстрее, чем при первичном.
У растений встречается еще один тип защитных реакций, индуцируемых патогенами, который называют индуцируемой системной устойчивостью (ИСУ). Этот тип защиты растительных организмов наблюдается, в частности, при инокуляции отдельными штаммами ризобактерий. ИСУ не требует для своей реализации салициловой кислоты, а регулируется такими фитогормонами, как жасмоновая кислота и этилен. ИСУ эффективна против тех патогенов, которые индуцируют жасмонат- и этилен-зависимые защитные механизмы.
Устойчивость растений к фитофагам. Вещества, служащие растениям для защиты от насекомых, очень разнообразны. Среди них имеются яды, способные убить любое животное. Поэтому животные обычно избегают ядовитых растений. Другие вторичные соединения придают растениям неприятный вкус, что также отпугивает травоядных животных. Для животных, как правило, также токсичны присутствующие в растениях многие терпеноиды и эфирные масла, цианогенные гликозиды и некоторые непротеиногенные аминокислоты. Цианогенные гликозиды при повреждении тканей разлагаются с выделением цианистой кислоты, а токсичность непротеиногенных аминокислот связана с тем, что они могут включаться в белки вместо обычных аминокислот и, таким образом, повреждать их.
В данной главе рассмотрена важная экологическая проблема – выявление закономерностей проявления невосприимчивости и путей практического использования этого свойства при создании устойчивых сортов сельскохозяйственных культур. Описаны общие закономерности и классификация явлений иммунитета растений к вредителям. Дан анализ имеющейся в литературных источниках информации по данной проблеме.
1.1. Наука об иммунитете: предмет, задачи, история развития
1.2. Устойчивость растений к вредителям
1.2.1. Биохимические особенности
Выбор вредителями растения для питания или наоборот, исключение из числа кормовых, основан на их способности улавливать биохимические различия разных видов и сортов.
Пример, колорадский жук предпочитает табак, картофель дикий и культурный. Биологические требования к составу пищи у насекомых часто меняются. По мере развития растения происходит смена хозяина.
1.2.2. Анатомо-морфологические особенности
Характер строения покровных тканей и анатомо-морфологические особенности растений влияют на избирательность вредителя.
Например, опушенность листьев мягкой пшеницы, более глубокие бороздки стимулируют яйцекладку гессенской мухи, а у твердой листья более гладкие и привлекательность вредителя меньше.
Панцирные сорта подсолнечника защищают семена от повреждения. Отродившиеся гусеницы не способны прогрызть панцирный слой оболочки, расположенный между пробковой и склеренхимной тканями. Это препятствует проникновению их в семянку.
1.2.3. Фенологические особенности
Многие вредители повреждают (или заселяют) растения только в определенные фазы их развития. Устойчивыми будут те, которые способны уйти от повреждений в критические фазы, когда потери ощутимы (шведская муха, просяной комарик и т. д.).
Устойчивые растения оказывают также антибиотическое действие на вредителя в момент питания и способны подавлять массовые развития вредителя либо изменять продолжительность их развития. Антибиотические действия обусловлены содержанием в устойчивых формах растений биологически активных веществ.
1.3. Типы повреждений растений вредными насекомыми и их ответная реакция
Вредители наносят повреждения в процессе питания, а некоторые и в период откладки яиц. Тип повреждений, наблюдаемый при питании, обусловлен строением ротового аппарата и биологическими свойствами вида. Он специфичен. Характер и степень повреждений растений определяют дальнейшую их жизнеспособность.
У вредителей с грызущим ротовым аппаратом различают следующие типы повреждений: скелетирования, сплошное объедание листьев и т. д.
Повреждения, вызываемые вредителями с колюще-сосущим ротовым аппаратом существенно отличаются от повреждений насекомых с грызущим ротовым аппаратом. Их внешний вид определяется характером ответных реакций растения на внедрение частей ротового аппарата и действия вводимых слюнных секретов. Различают два типа его внедрения в ткани растений.
Интрацеллюлярный тип — введение частей ротового аппарата в межклеточное пространство через естественные ходы без нарушения целостности клеточных стенок. Характерно для тлей.
Интрацеллюлозный тип — введение ротового аппарата в ткани растений путем прокалывания клеточных стенок. Характерно для клопов, цикадок, клещей.
Некоторым видам тлей свойственны оба типа внедрения. Вокруг мест питания вредители с колюще-сосущим ротовым аппаратом наблюдаются следующие нарушения: окраски, образование вздутий, галлов, усыхания, опадание цветов, бутонов и т. д.
Вредители с колюще-сосущим ротовым аппаратом вступают в более тесные взаимоотношения с поврежденным растением. Введя со слюной ферменты в растение, они способны к внеклеточному перевариванию тканей хозяина. У растения нарушается обмен веществ, в собственных ферментах усиливаются гидролитические процессы.
1.4. Типы устойчивости
Иммунитет растений к вредителям равно как и к микроорганизмам (бактерии, вирусы, грибы) является важным биологическим свойством обеспечивающим выживаемость растительных органов.
Существует большое разнообразие форм проявления устойчивости растений к вредителям, причем многие из них дополняют друг друга.
Различают три основных типа проявления устойчивости:
1) отвержение и выбор растений вредителями при откладке яиц или при питании (полная неповрежденность отдельных видов или сортов) растений определяется видом вредителей;
2) антибиотическое воздействие кормового растения на вредителя (неповреждаемость или слабая повреждаемость);
3) выносливость растений к повреждениям (см. гл. 2).
1.4.1. Отвержение и выбор растений вредителями
1.4.2. Антибиотическое воздействие кормового растения на вредителей
Антибиоз – это неблагоприятное воздействие растения на фитофага, проявляющееся при использовании его насекомым в пищу или для откладки яиц. В качестве факторов антибиоза могут выступать:
– вещества вторичного обмена;
– структурные особенности основных биополимеров, синтезируемых растениями и степень их доступности для усвоения фитофагами;
– пищевая ценность растения для вредителя;
– анатомо-физиологические особенности растений, затрудняющие доступ фитофага к зонам его наиболее благоприятного питания;
– ростовые процессы растений, приводящие к самоочищению растения от вредителя или нарушающие условия нормального развития фитофага.
На территориях, занятых устойчивыми сортами растений, обычно создаются условия, предотвращающие массовые размножения насекомых. Сорта, обладающие свойствами антибиоза, становятся мощным рычагом управления численностью насекомых.
1.5. Вещества вторичного обмена
Растениям наряду с синтезом веществ первичного обмена свойственен синтез веществ вторичного обмена, одной из функций которых является охрана целостности организма.
Вещества вторичного обмена представлены в виде различных эфирных масел, алколоидов гликозидов и других веществ, специфичных для определенных групп растений на уровне семейств, родов, а то и видов.
Например, колорадский жук перешел с дикорастущих пасленовых на картофель, который от своих диких сородичей отличается низким содержание гликоалколоидов, что улучшало питание и размножение вредителя. Устранение фактора ингибирующего размножение, обеспечили этому виду колоссальные возможности для захвата огромных территорий.
Эти вещества, безусловно, играли большую роль в историческом прошлом, являясь одной из причин становления пищевой специализации фитофагов (группы потребителей крестоцветных, пасленовых, маревых и т. д.).
Вредители капусты, например, приобрели способность к обезвреживанию ядовитого гликозида синергина и к использованию его в качестве источника энергии.
Вредители табака приспособились к никотину, вредители клещевины – к цианогену. Никотин, анабазин, пиретрин широко известны как инсектициды.
Адоптации фитофагов к использованию растений, содержащих те или иные вещества вторичного обмена относительны, поэтому при питании растениями, содержащими более высокие концентрации веществ вторичного обмена фитофаги начинают испытывать их отрицательное антибиотическое действие.
Наиболее чувствительны к веществам вторичного обмена личинки младших возрастов. Они гибнут уже в первые дни питания на 70–80 % (колорадский жук, гусеницы кукурузного мотылька).
Кроме гибели они могут вызывать расстройство пищеварительной, нервной, половой и других систем. В некоторых случаях отрицательное влияние этих веществ отмечено на отложенные яйца, на растения устойчивых сортов (колорадский жук, пьявица).
В селекции растений на устойчивость к вредителям оценка уровня содержания веществ вторичного обмена в различных сортах может усиленно использоваться в качестве маркера их устойчивости. Но вести селекцию на повышение концентрации этих веществ нельзя.
1.6. Структура и особенности основных биополимеров, синтезируемых растениями
Известно, что наиболее эффективным источником биологической энергии являются углеводы. При усвоении биополимеров растений в процессе пищеварения большое значение имеет степень стереохимического соответствия гидролитических ферментов насекомых молекулярным структурам пищи. При таком соответствии обеспечивается быстрый с минимальными затратами энергии гидролиз биополимеров и их всасывание. И наоборот, энергетические затраты на переваривание пищи насекомым возрастают при недостаточном стереохимическом соответствии между гидролизами и структурными особенностями биополимеров пищи.
Важное значение имеет также степень сбалансированности различных питательных веществ в соответствии с требованиями насекомых. Эта несбалансированность приводит к неэффективному использованию элементов пищи. Поэтому большое иммунологическое значение имеет уровень атакуемости биополимеров пищи гидролизами потребителя. При низком уровне атакуемости даже те растения, которые характеризуются высокой биологической ценностью, не могут быть использованы потребителем полностью. В результате этого у насекомых возникает так называемый синдром неполного голодания.
Усложнение формы полимеров пищевого субстрата свойственного устойчивым сортам приводит к различного рода отклонениям в жизнедеятельности фитофагов – усиливается секреция гидролитических ферментов, повышается их активность (клоп-черепашка, личинки злаковых мух и т. д.). При питании фитофагов на несвойственных для него тканях на устойчивых сортах создается дисбаланс в организме, что приводит к задержке в росте, снижению накопления жировых запасов, низкой подвижности, пониженной устойчивости к неблагоприятным условиям среды (при перезимовке) и часто к гибели.
Пищевые взаимоотношения возникли на основе многообразия типов питания и обмена веществ. Вначале они носили характер сосуществования между организациями, составляющими ценозы. Дальнейшая эволюция органического мира сопровождалась возникновением новых типов межвидовых отношений – появились бинарные и полинарные формы: симбиоз и паразитизм.
Система растение – насекомое—фитофаг относится к сложным. Организмы, составляющие эту систему, находятся на разных ступенях эволюционной лестницы, т. е. существенно различаются по уровню организации и, следовательно, наделены разными возможностями для реализации взаимодействия. В силу подвижности насекомых связи их с растениями в онтогенезе непостоянны во времени и для свободноживущих ограничиваются в основном актом еды или моментом откладки яиц. Хорошо развитые рецепторы обеспечивают широкие возможности выбора насекомыми кормового растения, различных его органов и тканей.
Малые размеры тела при общей высокой организации, высокая теплоотдача, подвижность в сочетании с огромным воспроизводительным потенциалом и другие специфические особенности делают насекомых весьма требовательными к пластическому и энергетическому обеспечению. Эта особенность насекомых превратила их в один из главных преобразователей энергии и информации в экосистемах как потребителей первичной и вторичной биологической продукции.
Общий характер отношений между насекомыми и растениями определяется двоякой ролью растений. Растение, выступая как компонент биоценоза, может играть роль внешнего фактора по отношению к насекомым-фитофагам, и в то же время, поступая в организм насекомого в качестве пищи, растение начинает играть роль и внутреннего фактора.


Рис. 1. A — золотарник высочайший (Solidago altissima). B и C — личинка и имаго питающегося на нем золотарникового листоеда (Trirhabda virgata). Изображение из статьи A. Kalske et al., 2019. Insect Herbivory Selects for Volatile-Mediated Plant-Plant Communication
Одна из наиболее явных причин такого переменного во времени и пространстве отбора для популяций растений — сукцессии, то есть закономерные смены сообществ на одной территории (например, смена березняка ельником или зарастание болота). Обычно этот процесс изучается на уровне реакции целого сообщества, но Айно Кальске (Aino Kalske) и Андре Кесслер (André Kessler) из Корнеллского университета (США) решили посмотреть, как популяции одного и того же вида реагируют на изменения пресса со стороны фитофагов на разных стадиях развития травянистого сообщества.
В качестве объекта они выбрали золотарник высочайший (Solidago altissima) (рис. 1, А) — многолетнее сложноцветное, популяции которого живут на заброшенных полях на протяжении десятилетий, переживая изменения в составе окружающей растительности и пресса фитофагов. Кстати, активно разрастающийся аналогичных местообитаниях в России золотарник канадский (Solidago canadensis) иногда рассматривается как синоним этого полиморфного вида.
Согласно гипотезе авторов, слабый пресс со стороны фитофагов, наблюдающийся в этом сообществе на ранних стадиях сукцессии (M. Howard et al., 2018. Eco-evolutionary processes affecting plant–herbivore interactions during early community succession), будет способствовать отбору генотипов с низким уровнем постоянной и индуцированной защиты, затем, на более поздних стадиях, из-за усиления активности фитофагов преимущество перейдет к генотипам с высоким уровнем индуцированной защиты, а если атаки фитофагов не прекратятся и станут более постоянными и предсказуемыми, то отбор будет поддерживать генотипы с высоким уровнем постоянной защиты.
Хотя золотарником питаются более сотни видов членистоногих, наибольший ущерб в месте исследования ему наносят жуки-листоеды Trirhabda virgata (рис. 1, В и С) и Microrhopala vittata. Ранее было показано, что устойчивость к ним наследуется и может эволюционировать в течение одного десятилетия: популяции, не сталкивающиеся с этими фитафагами, теряют устойчивые генотипы (R. F. Bode, A. Kessler, 2012. Herbivore pressure on goldenrod (Solidago altissima L., Asteraceae): its effects on herbivore resistance and vegetative reproduction). Основной способ индуцированной защиты золотарника — выработка в листьях фенольных соединений и дитерпеновых кислот.

Рис. 2. Аэрофотография экспериментального поля, разделенного на участки, находящиеся на разных стадиях девятилетней последовательности: три года засевания кукурузой, и шесть лет зарастания сорной растительностью, в том числе золотарником высочайшим. Два участка (late succession) не засевались 14 и 15 лет. Изображение из дополнительных материалов к обсуждаемой статье в Ecology
В 2017 году исследователи в течение всего сезона развития золотарника каждые три недели учитывали обилие двенадцати видов фитофагов, среди которых были и поедатели листьев, и листовые и стеблевые минеры (прокладывающие внутри тканей растений длинные ходы — мины), и галлобразователи (см. Галл).
Чтобы изучить уровень защиты от фитофагов в золотарниках из разных стадий сукцессии, исследователи собрали от 10 до 15 особей (под особью они понимали совокупность рамет — генетически идентичных стеблей, связанных общим корневищем) на участках, которые претерпели 1, 2, 4, 6 и 14 лет сукцессии после прекращения возделывания кукурузы. Выкопанные золотарники были перенесены в оранжерею, где их корневища дважды дорастили до новых побегов, чтобы избавиться от возможного влияния фитофагов, с которыми растения контактировали в поле.
Анализ сообществ фитофагов показал, что в первые годы сукцессии их состав существенно различается от участка к участку, но к 5–6 году смещается в сторону преобладания наиболее активных потребителей золотарника в регионе, T. virgata и M. vittata (рис. 3). Два участка с 15-летней сукцессией оказались сходны между собой и похожи на часть участков с ранних стадий сукцессии, обилие вышеупомянутых жуков на них также снизилось.
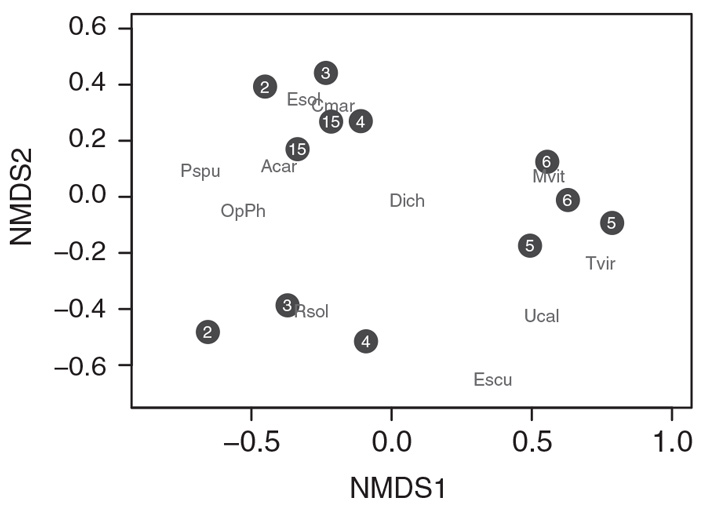
Рис. 3. Сообщества фитофагов на золотарниках, сходство которых изображено с помощью неметрического многомерного шкалирования: чем ближе друг к другу лежат черные кружочки с числами (обозначающими, сколько лет прошло с начала сукцессии), тем более похож состав фитофагов. Буквенные обозначения изображают обилие отдельных видов фитофагов в этом пространстве. Видно, что участки возрастом 5–6 лет похожи друг на друга и в основном населены M. vittata и T. virgata. Обозначения фитофагов: Acar — муха-галлица Asteromyia carbonifera, Cmar — клоп-кружевница Corythucha marmorata, Dich — выемчатокрылые моли Dichomeris spp., Escu — моль-листовертка Epiblema scudderiana, Esol — муха-пестрокрылка Eurosta solidaginis, Mvit — жук-листоед Microrhopala vittata, OpPh — минирующие мухи Ophiomyia sp. и Phytomyza sp., Pspu — пенница Philaenus spumarius, Rsol — муха-галлица Rhopalomyia solidaginis, Tvir — жук-листоед Trirhabda virgata, Ucal — тля Uroleucon caligatum. Рисунок из обсуждаемой статьи в Ecology
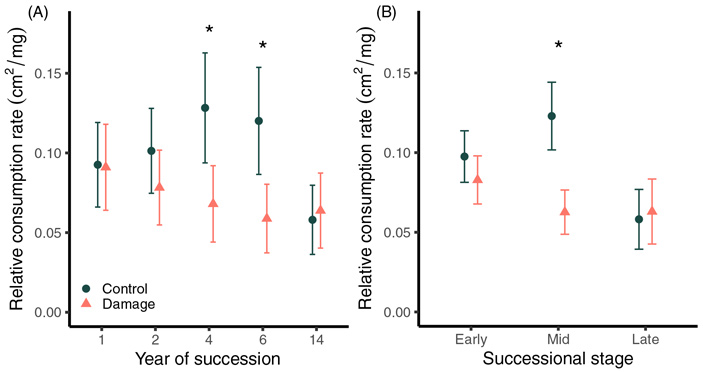
Рис. 4. Относительная скорость потребления золотарниковыми листоедами листвы золотарника высочайшего, взятого с разных стадий сукцессии и подвергшегося (damage) или не подвергшегося (control) предварительно поеданию этим же видом фитофага. Обратите внимание, что скорость потребления обратно зависит от устойчивости золотарника: чем ниже значение по вертикальной оси, тем более устойчив золотарник к поеданию. Звездочками отмечены значимые различия между подвергшимися и не подвергшимися атакам фитофагов золотарниками, свидетельствующие о существовании индуцированной защиты. Возраст участков, с которых были взяты растения, обозначен или как год с начала сукцессии (А), или как стадия сукцессии (В): ранняя, средняя и поздняя. Рисунок из обсуждаемой статьи в Ecology
Также обнаружилось, что скорость роста жуков отрицательно коррелировала с содержанием в листе производных кофейной и кумаровой кислот, но положительно — с содержанием флавоноидов, содержание же производных дитерпеновой кислоты никак не коррелировало с ростом жуков. При этом ни один из исследованных классов веществ не влиял на потребление личинками листьев.
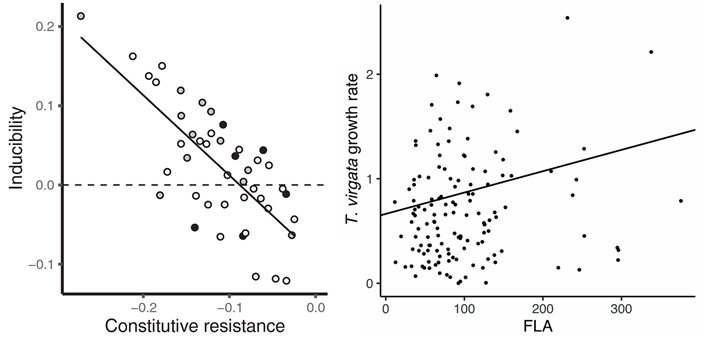
Наконец, в полном согласии с предыдущими исследованиями, постоянная и индуцированная защиты сильно отрицательно коррелировали друг с другом: у золотарников с высоким уровнем одной защиты был низкий уровень другой (рис. 5, слева).
Итак, исследование показало, что наиболее сильный пресс со стороны специализированных фитофагов золотарники испытывают в середине сукцессии, а не в начале и не в конце. Значит ли это, что популяция оказывается наименее устойчивой к поедателям спустя 4–6 лет после возникновения? Скорее всего, ситуация сложнее.
Авторы предполагают следующий сценарий, объясняющий это несоответствие. Где-то после пятого года золотарник перестает возобновляться семенами, так что далее на каждом конкретном участке существуют только определенные клоны. И высокий уровень поедания в средней сукцессии способствовал более активному разрастанию генотипов с высоким уровнем постоянной защиты. Поэтому к поздней сукцессии, когда низкий уровень поедания мог бы снова благоприятствовать индуцированной защите, таких генотипов уже не осталось.
Некоторые из полученных результатов, впрочем, выглядят контринтуитивно. Так, золотарниковый листоед лучше растет на листьях с высоким уровнем флавоноидов, которые вроде как должны защищать от поедания. Авторы предполагают, что дело в том, что T. virgata специализируется на питании растениями этого рода, что сделало его менее чувствительным к защитным веществам своей жертвы (H. V. Cornell, B. A. Hawkins, 2003. Herbivore responses to plant secondary compounds: a test of phytochemical coevolution theory). Впрочем, обнаруженная зависимость не отличается высокой статистической значимостью (р = 0,039), а ее график заставляет предположить, что положительный эффект возникает из-за двух точек в левом верхнем углу (рис. 5, справа).
Обсуждаемая работа не смогла раскрыть всех механизмов, влияющих на изменение проявляемой растениями устойчивости к фитофагам на разных стадиях сукцессии, но смогла показать, что свою роль в этом играет и отбор генотипов с определенной стратегией (в данном случае — с постоянной, а не индуцированной защитой), дающий эффект всего за несколько лет.

Растения противостоят инфекциям и другим стрессовым воздействиям окружающей среды посредством развития комплекса молекулярно-биохимических реакций (Яруллина, Ибрагимов, 2006). Восприятие, передача информации о действии факторов окружающей среды на растения и, соответственно, индукция ответных реакций, осуществляется с участием сигнальных систем (Тарчевский, 2002). Наиболее ранней ответной реакцией растительного организма на внедрение патогена является локальная генерация активных форм кислорода (АФК) - окислительный взрыв, запускающий цепь последующих защитных реакций (Аверьянов и др., 2007). Большую роль в генерации и регуляции содержания АФК в растительных тканях играют окислительно-восстановительные ферменты, в первую очередь, оксиредуктазы (Huckelhoven, Kogel, 2003). Показано, что образование АФК в ходе окислительного взрыва происходит при участии ферментов, локализованных на поверхности клеток: НАДФН-оксидаз, ксантиноксидаз, супероксиддисмутаз, оксалатоксидаз, пероксидаз (Gil-ad et al., 2000).
До недавнего времени считалось, что основным источником АФК при инфицировании являются только растительная ткань, а возможный вклад микроорганизмов в этот процесс не учитывался. Однако полученные в последние годы экспериментальные сведения ставят данное утверждение под сомнение (Гесслер и др, 2007). В частности, исследования, Аверьянова с соавторами (2006) показали, что скорость продукции АФК на листьях здоровых растений риса очень низка, но она значительно возрастает при инфицировании их возбудителем пирикуляриоза Magnaporthe grisea. Оказалось, что возбудитель пирикуляриоза риса способен к продукции АФК и в культуральной среде, вне растительного организма. Эти факты позволяют предположить, что патогенные грибы принимают активное участие в процессах продукции АФК. По-видимому, грибные антиоксиданты или супрессоры генерации АФК играют существенную роль в последующем формировании совместимых отношений в системе патоген - хозяин. Особая роль в этих процессах принадлежит ферментам патогенов: супероксиддисмутазам, каталазам, активация которых приводит к снижению токсичных для микроорганизмов концентрации АФК. Вероятно, локальная генерация АФК выполняет защитную функцию только на ранних этапах взаимодействия растения и патогена. Как известно, продолжительная и избыточная продукция АФК, характерная для восприимчивых растений, способствует развитию и распространению патогенов (Shirasu, Schulze-Lefert , 2000).
Для снижения уровня АФК в растительных тканях включаются механизмы детоксикации, связанные с активацией ферментов антиоксидантной системы. Возможно, что развитие или подавление следующих реакций в растительном организме, в первую очередь, зависит от концентрации Н2О2 в зоне контакта двух противоборствующих организмов.
Цель исследований наших состояла в выявлении ряда биохимических параметров, определяющих устойчивость растений и агрессивность патогенов при их взаимодействии. Объектами исследования были растения пшеницы (Triticum aestivum L.) и картофеля (Solanum tuberosum L), инфицированные грибными патогенами и насекомыми-вредителями.
Эффективным подходом к познанию механизмов развития устойчивости растений является сравнительное изучение реакций растений одного сорта на инфицирование штаммами патогена различной степени агрессивности. В соответствии с этим подходом, в экспериментах были использованы различные по агрессивности штаммы возбудителя септориоза злаковых - патогенного гриба Septoria nodorum. В предварительных опытах было показано, что штамм 9МН проявлял высокую, а штамм 4ВД - низкую агрессивность к исследуемым сортам яровой мягкой пшеницы (Жница, Омская 9, Саратовская 29, Башкирская 24). Штамм 9МН характеризовался более быстрым развитием в растительных тканях, более коротким латентным периодом культивирования и обильным спороношением по сравнению со штаммом 4 ВД. Оказалось, что инфицирование растений пшеницы восприимчивого сорта Жница высоковирулентным штаммом приводило к повышению интенсивности генерации АФК лишь в течение первых суток после инокуляции. Тогда как в инфицированных слабовирулентным штаммом тканях этого сорта, интенсивная продукция перекиси водорода происходила в течение более продолжительного периода времени (табл. 1). Вероятно, низкая концентрация Н2О2 не способна индуцировать развитие защитного ответами, и одновременно, является стимулирующей для роста патогена. Для снижения уровня АФК растения и грибы используют различные механизмы детоксикации, один из которых связан с активацией ферментов антиоксидантной системы (Белозерская, Гесслер, 2006). Показано, что активность каталазы в мицелии высоковирулентного штамма возбудителя мучнистой росы значительно выше, чем у низкоковирулентного (Талиева, Мишина, 1996). Ранее было показано, что высокий уровень генерации Н2О2, наблюдаемый в устойчивых растениях, способствует торможению активного роста и развития патогена, а низкий - инициирует его рост (Трошина, 2007).
Таблица 1.Содержание Н2О2 в листьях пшеницы сорта Жница при инфицировании грибом S. nodorum различной степени агрессивности
Читайте также: