
Показатели фотосинтетической продуктивности посева
Обновлено: 05.10.2024
В статье представлен процесс формирования фотосинтетического аппарата яровой пшеницы при применении биологических препаратов. Исследованиями установлено, что интенсивность нарастания листовой поверхности в течение всей вегетации достигала максимальных значений при инокуляции семян бактофосфином в смеси с ЖУСС-2 (жидкий удобрительно-стимулирующий состав) на фоне минеральных удобрений. При применении биопрепаратов интенсивность нарастания фотосинтетического потенциала посевов в течение вегетации яровой пшеницы возрастает за счет увеличения листовой поверхности. На характер формирования листовой поверхности яровой пшеницы минеральные удобрения и биопрепараты оказали непосредственное воздействие. Максимальные величины чистой продуктивности фотосинтеза наблюдались в фазе колошения, т.е. в период наибольшего развития листовой поверхности растений яровой пшеницы. Инокуляция семян способствует увеличению фотосинтетической деятельности растений в посевах и улучшению динамики ростовых процессов. Исследованиями установлено, что на характер формирования листовой поверхности яровой пшеницы минеральные удобрения и биопрепараты оказали непосредственное воздействие. Инокуляция семян биопрепаратами увеличивает данный показатель фотосинтетической деятельности, вероятно, за счет стимулирующего действия микроорганизмов, входящих в состав биопрепаратов, которая начинает проявляться с фазы кущения. Как при применении минеральных удобрений, так и без них, наибольшую площадь листьев имели растения на варианте бактофосфин + ЖУСС. Инокуляция семян способствует увеличению фотосинтетического потенциала посевов в течение вегетации за счет увеличения листовой поверхности. Инокуляция на фоне минеральных удобрений дает возможность формирования более высокого фотосинтетического потенциала посевов яровой пшеницы. При инокуляции семян яровой пшеницы биопрепаратами и препаратом ЖУСС-2 проявляется тенденция к повышению чистой продуктивности фотосинтеза как на неудобренном фоне, так и на фоне минеральных удобрений.

3. Мокроносов А.Т. Фотосинтетическая функция и целостность растительного организма [42-е Тимирязевские чтения]. – М.: Наука, 1983. – 63 с.
4. Мокроносов А.Т. Фотосинтез и продукционный процесс / А.Т. Мокроносов // Физиология растений на службе продовольственной программы СССР. – М.: Знание, 1988. – № 2. – С. 3–18.
6. Никитин С.Н. Влияние бактериальных удобрений на микрофлору почвы и урожайность яровой пшеницы // Агроэкологические аспекты повышения эффективности сельскохозяйственного производства: тез. докл. научно-практической конференции. – Пенза, 2001. – С. 127–129.
7. Ничипорович А.А. Некоторые принципы комплексной оптимизации фотосинтетической деятельности и продуктивности растений // Важнейшие проблемы фотосинтеза в растениеводстве. – М.: Колос, 1970. – С. 120–127.
8. Ничипорович А.А. Реализация регуляторной функции света и жизнедеятельности растений как целого и в его продуктивности. / А.А. Ничипорович. Фоторегуляция метаболизма и морфогенеза растений. – М., 1975. – 275 с.
9. Ничипорович А.А. Физиология фотосинтеза и продуктивность растений / А.А. Ничипорович. Физиология фотосинтеза. – М., 1982. – С. 7–34.
11. Ничипорович А.А. Фотосинтетическая деятельность растений в посевах / А.А. Ничипорович, Л.Е. Строганова, С.Н. Чмора, М.Н. Власова. – М.: Изд-во АН СССР, 1961. – 137 с.
12. Оценка эффективности микробных препаратов в земледелии / Под общ. ред. А.А. Завалина. – М.: РАСХН, 2000. – 82 с.
13. Федин М.А. Методика государственного сортоиспытания сельскохозяйственных культур / М.А. Федин. – М.: Изд-во Министерство сельского хозяйства СССР, 1985. – 285 с.
14. Шевелуха В.С. Периодичность роста сельскохозяйственных культур и пути ее регулирования. – М.: Колос, 1980. – 455 с.
Взаимосвязь растений в агроценозе носит непостоянный характер, зависящий от многих факторов. Главной задачей для получения высоких урожаев является создание такого посева, в котором бы максимально раскрывались потенциальные возможности фотосинтетической деятельности растений в агроценозе. Этого можно добиться при создании благоприятных условий для роста и развития растений.
Ряд авторов [3, 14] рассматривает рост растений как процесс дифференцирования организма за счет образования новых и увеличения старых элементов его структуры (молекул, клеток, тканей и органов), оказывающий решающее влияние на распределение, перераспределение и использование образовавшихся при фотосинтезе и метаболизме органических веществ, а также поглощение минеральных солей и воды, идущих на образование поверхности органов и тканей, их регенерацию и на запасные отложения. Рост растения суммирует в себе и венчает многие другие процессы жизнедеятельности растения, выражая в известной мере баланс процессов синтеза и распада веществ в организме при его взаимодействии с условиями внешней среды.
Рост растения и его биологическая продуктивность – результат, прежде всего, фотосинтетической деятельности, в ходе которой образуется до 95 % органических соединений. Поэтому рост растения, формативный, органообразовательный и рост как увеличение сухой биомассы начинается, главным образом, вслед за формированием фотосинтетической системы листа и осуществления процесса фотосинтеза. Лист как орган фотосинтеза является центром образования первичных продуктов, их метаболизации и эвакуации в органы запаса [2, 15].
Целью данной работы являлось выявление особенностей формирования фотосинтетической деятельности растений в посевах и динамика ростовых процессов при применении биологических препаратов и минеральных удобрений.
Материалы и методы исследования
Проводили следующие учеты и анализы:
– фенологические наблюдения согласно методике государственного сортоиспытания [13];
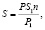
где S – общая площадь листьев одного растения, см2; S1 – площадь одной высечки, см2; P – общая масса листьев одного растения, г; P1 – масса высечек, г; n – число высечек. Зная густоту стояния растений на 1 га рассчитывалась площадь листьев на 1 га;
– содержание сухого вещества определялось отбором средней пробы измельченных растений. Из каждого образца отбирались навески по 50 г в двукратной повторности, фиксировались и высушивались в сушильном шкафу при температуре 105 °С до постоянной массы. Содержание сухого вещества определялось по формуле
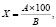
где А – масса навески после высушивания, г; В – масса навески до высушивания, г;
– чистая продуктивность фотосинтеза вычислялась по формуле Кидда, Веста и Бриггса:
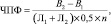
где ЧПФ – чистая продуктивность фотосинтеза; В1 и В2 – сухая биомасса пробы урожая в начале и конце учетного периода, г; В2 – В1 – прирост сухой массы за учетный период n дней, г; (Л1 + Л2)/0,5 – средняя работавшая площадь листьев за этот промежуток времени, м2; n – число дней.
Минеральные удобрения аммиачная селитра, двойной гранулированный суперфосфат и хлористый калий вносили под предпосевную культивацию. Обработка семян бактериальными препаратами и препаратом ЖУСС-2 (жидкий удобрительно-стимулирующий состав) проводили за 1–2 дня до посева. Повторность опыта 4-кратная. Учетная площадь делянок 42 м2 (2,1*20).
Результаты исследования и их обсуждение
Формирование фотосинтетического аппарата представляет сложный процесс. В ранние фазы роста и развития преобладают процессы новообразования и роста листьев, а в более поздние – процессы отмирания, связанные с усиленной транспортировкой пластических веществ в репродуктивные органы [9]. Накопление и запасание энергии в процессе фотосинтеза сопровождается накоплением биомассы, служащей структурным и энергетическим материалом, обеспечивающим существование растений [15].
Максимальные урожаи могут быть сформированы посевами с оптимальной площадью листьев, причем важно, чтобы она быстро нарастала до максимальной величины и долго удерживалась на достигнутом уровне без резкого снижения к концу вегетации, максимально поглощая солнечную радиацию [10]. У культур, потенциально способных к формированию очень большой листовой поверхности, к которым относятся, в частности, злаки [8], степень ее фактического развития, а тем самым и факторы фотосинтетической деятельности, лимитирующие урожайность, решающим образом зависят от условий выращивания. Если ресурсы влаги и питания недостаточны, то основным фактором, лимитирующим урожайность растений, является недостаточное развитие листовой поверхности, а на бедных почвах – и низкая производительность ее работы.
Основными показателями фотосинтетической деятельности растений являются: площадь листьев, фотосинтетический потенциал, чистая продуктивность фотосинтеза и коэффициент использования фотосинтетически активной радиации. Величина площади листьев слагается из площади листьев отдельных растений и в различной степени зависит от периода вегетации, погодных условий выращивания, сорта, густоты растений, обеспеченности их питательными веществами и т.д.
Исследования [4, 5] указывают, что более продуктивным может быть то растение, которое отличается менее интенсивным фотосинтезом, но больший процент ассимилятов использует на образование листьев и формирует большую ассимиляционную поверхность. Поэтому очень важно агротехническими мероприятиями в наиболее короткие сроки добиваться оптимальной площади листьев растений.
Имеется немало указаний в литературе, что с увеличением ассимиляционной площади листьев увеличивается урожай. Однако положительная связь этих двух процессов имеет предел, при котором большая листовая поверхность из-за взаимного затенения снижает интенсивность фотосинтеза, в результате чего увеличивается непродуктивная часть урожая и уменьшается продуктивная.
Наши исследования показывают, что в начале развития растений яровой пшеницы от появления всходов до фазы кущения только начинают проявляться различия вариантов по площади листьев. Площадь листьев в фазу кущения изменяется в пределах 7,5–8,9 тыс. м2/га, а на фоне минеральных удобрений 9,1–11,4 тыс. м2/га (табл. 1). При инокуляции семян наблюдается тенденция к увеличению ассимиляционной поверхности листьев, особенно это заметно при применении бактофосфин + ЖУСС-2. По-видимому, в данный период действие биопрепаратов только начинает проявляться [6].
Влияние биопрепаратов на ассимиляционную поверхность листьев, тыс. м2/га (среднее за 3 года)
Урожай создается в процессе фотосинтеза, когда в зеленых растениях образуется органическое вещество из диоксида углерода, воды и минеральных веществ. Энергия солнечного луча переходит в энергию растительной биомассы. Эффективность этого процесса и в конечном счете урожай зависят от функционирования посева как фотосинтезирующей системы.
В полевых условиях посев (ценоз) как совокупность растений на единице площади представляет собой сложную динамическую саморегулирующуюся фотосинтезирующую систему. Эта система включает в себя много компонентов, которые можно рассматривать как подсистемы; она динамическая, так как постоянно меняет свои параметры во времени; саморегулирующаяся, так как, несмотря на разнообразные воздействия, посев изменяет свои параметры определенным образом, поддерживая гомеостаз.
Такую систему характеризуют новые свойства по сравнению с отдельным растением. Так, для отдельного растения увеличение площади питания и связанное с этим улучшение освещенности приводят к повышению его семенной продуктивности, а для ценоза важна оптимальная густота растений. Условия для максимальной продуктивности отдельного растения и ценоза как системы не совпадают.
Исследованию фотосинтеза ценозов была посвящена Международная биологическая программа. Изучение фотосинтетической деятельности растений в посевах тесно связано с теорией получения высоких урожаев и возможностью управления формированием урожая. Методические основы ее изучения разрабатывали многие ученые. Большой вклад внес А. А. Ничипорович.
Управление формированием урожая весьма сложно, так как растения в ценозе, изменяясь в процессе вегетации, взаимодействуют с другими сложными системами — микроорганизмами почвы, возбудителями болезней, сорняками, вредителями. Многие факторы среды — температурный режим, осадки и др. — практически невозможно контролировать. Однако на основании анализа природно-климатических факторов можно подобрать сорта, адаптированные к конкретным условиям, разработать технологию их возделывания. Многие факторы поддаются регулированию. Можно изменить условия минерального питания, воздействовать на сорняки, вредителей и болезни. Управление процессом формирования урожая ведут на основе систематического контроля за развитием растений и направления хода фотосинтетической деятельности посевов в соответствии с заранее заданными параметрами.
Фотосинтетически активная радиация
Необходимое условие фотосинтеза — энергия солнечной радиации. Следует создавать такие посевы, в которых листья поглощали бы энергию солнца с возможно более высоким коэффициентом полезного действия для создания наибольшей биомассы и сосредоточения ее в хозяйственно ценной части урожая — семенах, клубнях, корнеплодах и т. п.
В процессе фотосинтеза принимает участие не вся солнечная энергия, а только ее видимая часть — фотосинтетически активная радиация (ФАР) с длиной волн от 380 до 720 нм (нанометр). Эти лучи поглощаются хлорофиллом и являются энергетической основой фотосинтеза. Энергия ФАР составляет около 50 % общей энергии солнечной радиации. Инфракрасная часть солнечного спектра, составляющая также около 50 % общей энергии солнца, не участвует в фотохимических реакциях фотосинтеза. Эти лучи поглощаются почвой, от которой нагреваются приземный слой воздуха и сами растения, при этом усиливаются транспирация и испарение влаги с поверхности почвы. Количество ФАР, падающее на единицу поверхности почвы в среднем по месяцам года и по декадам месяца, определено для различных географических зон и приведено в соответствующих справочниках.
Объективным показателем величины урожая (высокий, средний, низкий) может служить коэффициент использования ФАР. Хорошие урожаи соответствуют 2…3 % использования ФАР. При возделывании сортов интенсивного типа и оптимизации всех процессов формирования урожая возможна аккумуляция в урожае 3,5…5,0 % ФАР и более.
Поступление солнечной энергии за вегетационный период зависит от географической широты. Так, если сравнивать северные и южные районы России, то приход ФАР за возможный вегетационный период различается более чем в 2 раза. Кроме того, можно отметить, что на плодородных почвах Краснодарского края при достаточном количестве тепла и влаги значительно легче сформировать посев, который в среднем за вегетацию усваивает 3 % ФАР, чем в Тверской области на малоплодородных почвах и при недостатке тепла.
Показатели фотосинтетической деятельности посевов
Посев представляет собой оптическую систему, в которой листья поглощают ФАР. В начальный период развития растений ассимиляционная поверхность невелика и значительная часть ФАР проходит мимо листьев, не улавливается ими. С повышением площади листьев увеличивается и поглощение ими энергии солнца. Когда индекс листовой поверхности* составляет 4…5, т. е. площадь листьев в посеве 40…50 тыс. м2/га, поглощение ФАР листьями посева достигает максимального значения — 75…80 % видимой, 40 % общей радиации. При дальнейшем увеличении площади листьев поглощение ФАР не повышается.
В посевах, где ход формирования площади листьев оптимальный, поглощение ФАР может составить в среднем за вегетацию 50…60 % падающей радиации. Поглощенная растительным покровом ФАР — энергетическая основа для фотосинтеза. Однако в урожае аккумулируется только часть этой энергии. Коэффициент использования ФАР обычно определяют по отношению к падающей на растительный покров ФАР. Если в урожае биомассы в средней полосе России аккумулировано 2…3 % прихода на посев ФАР, то сухая масса всех органов растений составит 10… 15 т/га, а возможная урожайность — 4…6 т зерна с 1 га. В изреженных посевах коэффициент использования ФАР составляет всего 0,5…1,0%.
При рассмотрении посева как фотосинтезирующей системы урожай сухой биомассы, создаваемый за вегетационный период, или его прирост за определенный период зависит от величины средней площади листьев, продолжительности периода и чистой продуктивности фотосинтеза за этот период.
где У —урожайность сухой биомассы, т/га; ФП— фотосинтетический потенциал, тыс. м 2 • дни/га; ЧПФ —чистая продуктивность фотосинтеза, г/(м 2 • дни).
Фотосинтетический потенциал рассчитывают по формуле
где Sc — средняя за период площадь листьев, тыс. м 2 /га; Т — продолжительность периода, дни.
Основные показатели для ценоза, как и урожайность, определяют в расчете на единицу площади —1м 2 или 1 га. Так, площадь листьев измеряют в тыс. м 2 /га. Кроме того, пользуются таким показателем, как индекс листовой поверхности.
Динамика площади листьев в посеве подчиняется определенной закономерности. После появления всходов площадь листьев медленно повышается, затем темпы нарастания увеличиваются. К моменту прекращения образования боковых побегов и роста растений в высоту площадь листьев достигает максимальной за вегетацию величины, затем начинает постепенно снижаться в связи с пожелтением и отмиранием нижних листьев. К концу вегетации в посевах многих культур (зерновые, зерновые бобовые) зеленые листья на растениях отсутствуют.
Динамика площади листьев показывает, что на разных этапах вегетации посев как фотосинтезирующая система функционирует неодинаково (рис. 3). Первые 20…30 дней вегетации, когда средняя площадь листьев составляет 3…7 тыс. м2/га, большая часть ФАР не улавливается листьями, и поэтому коэффициент использования ФАР не может быть высоким. Далее площадь листьев начинает быстро нарастать, достигая максимума. Как правило, это происходит у мятликовых в фазе молочного состояния зерна, у зерновых бобовых в фазе полного налива семян в среднем ярусе, у многолетних трав в фазе цветения. Затем площадь листьев начинает быстро снижаться. В это время преобладают перераспределение и отток веществ из вегетативных органов в генеративные.

Рис. 3. Оптимальная площадь листьев в посевах однолетних культур с длительностью вегетационного периода 100 и 120 дней
На продолжительность этих периодов и их соотношение влияют различные факторы, в том числе агротехнические. С их помощью можно регулировать процесс нарастания площади листьев и продолжительность периодов. В засушливых условиях густоту растений, а следовательно, и площадь листьев намеренно снижают, так как при большой площади листьев усиливается транспирация, растения сильнее страдают от недостатка влаги, урожайность уменьшается.
Слишком большое разрастание площади листьев при достаточном водоснабжении также приводит к нежелательным результатам. Биомасса в этом случае растет довольно высокими темпами за счет вегетативных органов, однако условия формирования плодов и семян ухудшаются. К подобным результатам может привести и чрезмерное загущение растений. Для кормовых культур, у которых листья представляют хозяйственно ценную часть урожая (например, травы), площадь листьев может достигать 60…80 тыс. м 2 /га.
Таким образом, получение высоких урожаев связано с оптимальным ходом (графиком) нарастания площади листьев. Такие графики могут быть определены для каждой культуры и сорта в конкретных условиях их выращивания.
Формирование урожая зависит не только от величины площади листьев, но и от времени ее функционирования. Фотосинтетический потенциал (ФП) объединяет эти показатели. ФП может быть определен за любой период времени, например за декадные, межфазные периоды или в целом за вегетационный период. ФП за какой-либо период представляет сумму величин площади листьев за каждые сутки периода. Например, если площадь листьев в начале периода составляла 20 тыс. м 2 /га, а через 10 дней — 28 тыс. м 2 /га, то ФП этого 10-дневного периода составляет (20 + 28): 2 • 10 = = 240 тыс. м 2 • дни/га. ФП хорошо развитых посевов зерновых культур с вегетационным периодом 100…ПО дней составляет за вегетацию 2,0…2,5 млн м 2 • дни/га. В южных районах для культур с длительным вегетационным периодом и при хорошем водоснабжении ФП может составить до 4 млн м 2 • дни/га.
Чистая продуктивность фотосинтеза (ЧПФ) характеризует интенсивность фотосинтеза посева и представляет собой количество сухой массы растений в граммах, которое синтезирует 1 м 2 листовой поверхности за сутки. В среднем за вегетацию у таких культур, как пшеница, ячмень, ЧПФ составляет 5…7 г/(м 2 • дни). У кукурузы ЧПФ обычно выше. ЧПФ, так же как и ФП, определяют за какой-либо период или в среднем за вегетацию:
где В2 и B1 — сухая масса растений с единицы площади в конце и в начале периода.
ЧПФ варьирует в течение вегетации. В первый месяц вегетации ЧПФ выше, чем в последующий, так как в начале вегетации растения не затеняют друг друга, все листья хорошо освещены. В дальнейшем с увеличением площади листьев ЧПФ начинает уменьшаться в связи с затенением нижних листьев.
В начале вегетации нарастание биомассы идет медленно, затем темпы приростов увеличиваются. В конце вегетации, когда площадь листьев небольшая, суточные приросты биомассы также невелики. В это время идет перераспределение накопленных ассимилятов из листьев, стеблей и корней в генеративные органы.
Прирост биомассы за любой промежуток времени, в том числе и за вегетацию, равен произведению ФП и ЧПФ. Если в среднем за 100 дней вегетации ЧПФ равнялась 6 г/(м 2 • дни), а ФП — 2 млн м 2 • дни/га, то количество сухой биомассы составит 12 т/га.
Посев как фотосинтезирующая система наиболее производительно функционирует в период, когда площадь листьев близка к оптимальной — 30…50 тыс. м 2 /га. Если ЧПФ в это время равна 5…7 г/(м 2 •дни), то при площади листьев 40 тыс. м 2 /га суточный прирост сухой биомассы составит 200…280 кг/га. Если период с такой средней площадью листьев продолжается 30 дней, то за это время прирост сухой биомассы составит 6…8 т/га. Прирост биомассы за период, когда посев функционирует в оптимальном режиме, составляет более 70 % максимального за вегетацию, хотя продолжительность этого периода — всего 30 % общей вегетации культуры. Коэффициент использования ФАР в это время в 2…3 раза больше, чем в первый месяц после появления всходов, а также в конце вегетации.
При созревании в корнях и стеблях сосредотачивается 50…60 % сухой массы растений, в основном представленной клетчаткой. В нашем примере из общей биомассы 12 т/га на урожай зерна приходится 5…6 т.
Различия в урожаях в конечном итоге определяются тем, достигал ли и как скоро достигал посев в своем развитии оптимального для данных условий состояния, а также как долго он функционировал в этом состоянии.
Таким образом, высокие урожаи обеспечиваются определенным ходом фотосинтетической деятельности растений в посевах. Оптимальный ход нарастания площади листьев и биомассы может быть определен для каждой культуры и сорта в конкретных условиях выращивания.
Факторы, лимитирующие фотосинтез
Почему растения в посеве не достигают оптимального уровня показателей фотосинтетической деятельности или существенно отклоняются от них в отдельные периоды вегетации? Какие элементы фотосинтетической деятельности подвержены наибольшему влиянию различных факторов?
Как следует из формулы У = ФП • ЧПФ, урожай биомассы (а также и хозяйственно ценной части урожая) прямо зависит от этих показателей. При улучшении условий жизни растений (оптимизация режима питания и влагообеспеченности) обычно усиливаются ростовые процессы, увеличивается площадь листьев. В этом случае листья сильнее затеняют друг друга, поэтому ЧПФ снижается.
Большинство культурных растений относятся к типу С3, у которых фотосинтез усиливается с ростом концентрации СO2 при насыщающей интенсивности освещения.
К растениям типа С4 относятся хлеба второй группы — кукуруза, сорго, просо, рис и сахарный тростник. У них не наблюдается светового насыщения и усиленного фотодыхания, а компенсационная точка по СO2 необычайно низка. ЧПФ у С4-растений выше, чем у С3-растений, особенно при повышенной площади листьев. Сорта детерминантного типа роста (ультраскороспелые сорта сои) обычно характеризуются более высокой ЧПФ. Имеют значение также форма, направление и анатомическое строение листьев. У сортов люпина узколистного ЧПФ обычно выше, чем у сортов других видов люпина.
Считается, что у зерновых культур фотосинтез посева осуществляется лучше, если верхние листья направлены под острым углом к стеблю. В процессе селекционного совершенствования сахарной свеклы распластанная по поверхности почвы розетка листьев постепенно превращалась в воронкообразную, что определило способность современных сортов создавать высокопродуктивные посевы.
Большое значение имеет аттрагирующая способность генеративных и запасающих органов растений, благодаря которой пластические вещества из листьев активно перемещаются в них.
Для скороспелых форм ФП должен составлять 1,5…2,0 млн м 2 • дни/га. для среднеспелых — 2,5…3,0 и для позднеспелых — 3…5 млн м 2 • дни/га.
В пределах одного типа скороспелости увеличение ФП может происходить путем ускорения темпов роста площади листьев и увеличения ее максимального значения. Густота посевов —один из факторов, сильно влияющих на эти показатели. Однако чрезмерное разрастание листьев при усилении их затенения может стать отрицательным фактором для формирования репродуктивных органов.
При оптимальной влагообеспеченности фактором, ограничивающим ростовые процессы, становится уровень минерального питания. Для получения высоких урожаев необходима высокая обеспеченность элементами минерального питания, в первую очередь азотом. Так, растения пшеницы при урожайности зерна 5 т/га выносят из почвы 170…200 кг азота с 1 га. Однако высокие нормы азота при достаточном количестве влаги усиливают вегетативный рост растений и разрастание листовой поверхности сверх оптимальных размеров, растения полегают, урожай снижается.
Основное средство повышения эффективности использования ценозом влаги и удобрений на фотосинтез и повышение урожайности — генетическое преобразование растений, создание сортов, у которых рост вегетативных органов ограничен, а генеративных — значительно выше, чем у старых сортов. Поэтому одно из направлений современной селекции — создание сортов, реагирующих на улучшение условий выращивания увеличением хозяйственно ценной части урожая.
В онтогенезе растений от всходов до начала созревания выделяют 4 биологически обоснованных периода:
- всходы — начало цветения;
- цветение и образование плодов;
- рост плодов;
- налив семян.
Основные выходные показатели первого периода — площадь листьев и ФП; второго — максимальная за вегетацию площадь листьев, ФП этого периода и количество образовавшихся плодов в расчете на единицу площади (на 1 м 2 ). Число образовавшихся плодов прямо коррелирует с ФП первого периода и особенно тесно — с ФП второго.
В третий период (рост плодов) площадь листьев постепенно начинает уменьшаться, но в среднем сохраняется на высоком уровне; биомасса продолжает интенсивно нарастать за счет роста плодов, которые к концу периода достигают максимальной величины. От числа сохранившихся к концу третьего периода плодов и семян зависят показатели фотосинтеза и поступление в растения азота в четвертый период, когда происходит налив семян. Посев как фотосинтезирующая система наиболее активно функционирует в течение второго и третьего периодов. За это время, равное по продолжительности первому периоду, накапливается 60…70 % биомассы и такое же количество азота от максимальных за вегетацию величин.
Таким образом, эффективность работы каждого последующего периода зависит не только от агрометеорологических условий этого периода, но и от результатов функционирования посева в предыдущий период. Конечные показатели развития посева — урожай семян и его качество — зависят от последовательных состояний посева в процессе роста и развития растений.
Величина площади листьев изменялась в зависимости от вида, дозы и комбинированного применения гербицидов, погодных условий и густоты стояния растений. Причем, темпы нарастания листовой поверхности были одинаковыми по вариантам. В среднем за годы исследований в фазу 2—3 настоящих листьев наиболее развитый листовой аппарат имели растения на контрольном варианте с трефланом (1,75 кг/га), за исключением вариантов с увеличенной нормой высева. Объясняется это тем, что эрадикан в дозе 4 кг/га и прометрин в дозе 2 кг/га оказывают несколько угнетающее действие на растения сои в начальный период роста. Площадь листовой поверхности на этих вариантах была соответственно на 13,2 и 9,6 % меньше контроля. Совместное применение эрадикана (4 кг/га) и прометрина (0,75 кг/га) еще больше снизило площадь листовой поверхности — на 16,9—21,7 %, по сравнению с контролем. Сочетание прометрина (0,75 кг/га) с трефланом (1,75 кг/га) вызвало менее заметное угнетение молодых растений сои, и площадь листьев была здесь, по сравнению с контролем, на 4,8—8,4 % меньше. На делянках с узкорядным посевом и увеличенной (500 тыс. шт/га) нормой высева семян площадь листьев была на 8,4—10,8 % больше, чем у контроля, и только за счет количества растений на единице площади (табл. 7.3, рис. 7.1).





На делянках с широкорядным посевом и нормой высева 400 тыс. шт/га, наименьшая зеленая и сухая массы были в этот период при использовании эрадикана в чистом виде и в смеси с прометрином — 17,1—18,6 ц/га и 3,7—4,1 ц/га, что на 20,1—13,1 % и 21,2—15,7 % меньше контроля. Среднесуточный прирост сухого вещества здесь был наименьшим за период вегетации — 17,2—17,8 кг/га, т. е. на 12,7—15,7 % меньше контроля. Объясняется это угнетающим действием препарата на растения сои в начальный период роста, меньшим количеством растений на единице площади и нерациональным размещением растений при данном способе посева. Несколько меньшее снижение зеленой и сухой массы (4,6—6,5 и 4,2—8,5 %), по сравнению с контролем, было на делянках с таким же способом посева и нормой высева обработанных перед посевом полей прометрином в чистом виде и в смеси с трефланом, что объясняется меньшим отрицательным влиянием гербицидов на растения сои.


Исследованиями А.А. Ничипоровича (1966) и А.Я. Бакалдина (1973) установлено, что урожай сухого вещества сельскохозяйственных культур на 80–90 % создается в результате фотосинтеза, который, в первую очередь, зависит от размеров ассимиляционной поверхности, высоты и густоты стояния растений и ряда других факторов. Все остальные процессы питания растений, в частности водное и минеральное, эффективны в той степени, когда они обеспечивают и поддерживают оптимальную деятельность фотосинтетического аппарата.
Нарастание сухой массы считается одним из главных показателей фотосинтетической деятельности растений. Суточный прирост сухого вещества значительно изменяется и может достигать до 300 кг/га в период интенсивного роста растений (А.А. Ничипорович и др., 1961).
Потенциальная биологическая продуктивность полевых культур зависит от величины поступающей к ним солнечной энергии и от особенностей её расхода. Помимо продуцирования кислорода, фотосинтез приобретает всё большее значение как основа воспроизводства продовольствия. Поэтому, создавая искусственные ценозы на полях, человек при помощи агротехники и лучистой энергии солнца обеспечивает себя продуктами питания. Первичные продукты фотосинтеза, участвуя в реакции вторичного метаболизма, образуют весь спектр органических соединений растительной клетки (А.Т. Мокроносов, В.Ф. Гавриленко, Т.В. Жигалова, 2006).
Высокая продуктивность сельскохозяйственных культур определяется динамически оптимальным соотношением отдельных элементов фотосинтеза. К основным из них относят: размер ассимиляционного аппарата, фотосинтетический потенциал, интенсивность и продуктивность фотосинтеза (Т.М. Русакова, 1974; J.M. Anderson, 1983; В.М. Важов, 2012).
Известно, что годовая масса органического вещества, создаваемого фотосинтезом, с избытком перекрывает то количество продовольствия, которое необходимо населению планеты. Поэтому одним из путей повышения эффективности земледелия является создание посевов с хорошей структурой пространственного расположения листьев.
Листовая поверхность является главным рабочим органом зелёных растений, а размер урожая почти полностью определяется её величиной. Нарастание листовой поверхности и величина ассимиляционного аппарата культурных растений зависит от агротехнических условий (А.Н. Бегишев,1953; Vong Hguyen Quoc, Murata Yoshio, 1978) .
По вопросу об оптимальной площади листовой поверхности сельскохозяйственных культур нет единого мнения. А.А. Ничипорович (1959) считает, что листовая поверхность должна составлять до 50 тыс. м2/га, а по мнению А.Н. Бегишева (1953) этот показатель равен примерно 100 тыс. м2/га.
В исследованиях А.Д. Гончарова (2008) в Новосибирской области бороздковые посевы формировали лучшую листовую поверхность гречихи, чем сплошные. С возрастом растений преимущество бороздковых посевов было очевидным. В фазу бутонизации площадь листьев на 1 га бороздковых посевов была больше рядовых на 14,5 %, а в фазу цветения – на 37,4 %. В изменении площади листьев по годам исследований каких-либо закономерностей не отмечено как по каждому способу возделывания, так и по сравнению их между собой. В различиях между изучаемыми способами посева по другим показателям фотосинтетической деятельности гречихи и их динамики в течение вегетационного периода в опытах А.Д. Гончарова (2008) проявились те же закономерности, что и в случае с площадью листьев. Так, фотосинтетический потенциал бороздковых посевов был больше соответствующего показателя сплошных посевов на 15 % за период бутонизация-цветение и на 30 % – за время цветение-созревание.
По мнению Н.Д. Кумсковой (2004), в условиях Дальнего Востока на листовую поверхность гречихи влияют сроки сева. В её опытах посевы 5 и 30 июня имели площадь листьев 22,5 и 23,5 тыс. м2 на 1 га, тогда как посевы 20 и 25 июня, соответственно, 32 и 31 тыс. м2. Гречиха давала хорошую вегетативную массу и высокие показатели фотосинтеза. В опыте Н.Д. Кумсковой (2004) при посеве с 5 по 30 июня получено от 25,1 до 37,3 т/га зелёной массы, максимальной она была при сроке сева 20 июня (табл. 56).
Фотосинтетическая деятельность и урожайность вегетативной массы гречихи
на разных сроках сева (по Н.Д. Кумсковой, 2004)
Площадь листьев (тыс. м2/га)
Зеленая масса, т/га
Сухая масса, т/га
Примечание. ЧПФ – чистая продуктивность фотосинтеза за вегетацию, г/м2/сут.
По мнению Н.Д. Кумсковой (2004), влияние удобрений на площадь листьев, продуктивность фотосинтеза, прирост сухого вещества гречихи значительное (рис. 61).
Прирост зелёной и сухой массы гречихи находился в тесной зависимости от удобрений. На вариантах без их применения и при внесении азота прирост зелёной массы продолжался до уборки, что важно при выращивании гречихи на зелёное удобрение. Внесение фосфора, совместно с комплексным удобрением, обеспечивало накопление биомассы на 1–2-е декады раньше (табл. 57).

Рис. 61. Влияние удобрений на рост площади листьев, продуктивность фотосинтеза, прирост сухого вещества гречихи по декадам (по Н.Д. Кумсковой, 2004)
Динамика нарастания зелёной и сухой массы гречихи в зависимости от удобрений (по Н.Д. Кумсковой, 2004), т/га
Удобрения, в зависимости от влажности почвы, по-разному влияли на формирование листовой поверхности гречихи (Н.Д. Кумскова, 2004). При выпадении большого количества осадков в начальный период вегетации на варианте с азотом формировалась низкая площадь листьев – 21,7 тыс. м2 на 1 га. На делянках, удобренных только фосфором и при совместном внесении азота, фосфора и калия листовая поверхность была больше на 19,3 и 25,1 тыс. м2 на 1 га, по сравнению с вариантом N40, и на 8,4 и 14,2 тыс. м2 – по отношению к контролю.
Существенно изменялась листовая поверхность в зависимости от густоты посева (Н.Д. Кумскова, 2004). С увеличением нормы высева с 2 до 5 млн. всх. зёрен на 1 га площадь листьев на одном растении снизилась с 175 до 104 см2, а на 1 га увеличилась с 26 до 57 тыс. м2.
Т.М. Русакова (1974) также считает, что площадь листьев гречихи на высоком агрофоне может превышать показатели контроля в 2,0–2,5 раза.
С.У. Броваренко (1970), на основании многолетних исследований в Западной Сибири, пришёл к выводу о том, что лучшее развитие ассимиляционной поверхности листьев гречихи происходит на узкорядных посевах, в сравнении с широкорядными. Площадь листьев на узкорядных посевах достигает 44,4 тыс. м2 на 1 га, на рядовых – 29,8, а на широкорядных – только 19,6 тыс. м2 на 1 га, соответственно, и урожайность на последних ниже. Можно высказать предположение, что урожайность на широкорядных посевах снижалась из-за угнетения культурных растений сорняками, так как известно, что рядовые, и особенно узкорядные посевы, успешно противодействуют засорённости.
С.И. Рак (1967) также считает, что площадь листьев гречихи при рядовом способе посева примерно в 1,5 раза больше в сравнении с листовой поверхностью на междурядьях 0,45 м.
По мнению К.А. Савицкого (1970), большое производственное значение имеют широкорядные посевы гречихи. На чистых от сорняков почвах они эффективнее сплошных рядовых. Листовая поверхность на широкорядном посеве в 1,3–1,4 раза больше, чем на сплошном. Это способствует повышению продуктивности фотосинтеза у растений широкорядного посева в фазе плодообразования в 1,5–1,9 раза.
Наши наблюдения подтверждают данные результаты. В зависимости от ширины междурядий площадь листьев возрастает в 1,4 раза в пользу разреженного посева, увеличение нормы высева также способствует росту ассимиляционной поверхности примерно в 1,5 раза.
Преимущество узкорядного способа посева перед обычным рядовым и широкорядным можно объяснить более слабым ростом, развитием и ветвлением растений в разреженных посевах, что наблюдается при наличии сорняков в междурядьях.
Гречиха интенсивно формирует листья в фазу всходов и бутонизации, к началу цветения их доля составляет 30–50 % от всей биомассы (Г.В. Копелькиевский, 1963). Способы посева и нормы высева значительно влияют на формирование площади листьев. Так, к периоду начала плодообразования размер листовой поверхности быстро увеличивается, но затем скорость данного процесса снижается.
Наши наблюдения показали, что в условиях лесостепи Алтайского края площадь листьев гречихи в фазу плодообразования составляет 48,9–69,8 тыс. м2/га (табл. 58).
Площадь листьев и продуктивность фотосинтеза посевов гречихи
(средняя за 2010–2011 гг.)
Норма высева, млн. всх. зёрен на 1 га
площадь листьев, тыс. м2/га
площадь листьев, тыс. м2/га
площадь листьев, тыс. м2/га
Широкоряд-ный (0,45 м)
Широкоряд-ный (0,60 м)
В связи с тем, что по урожайности зерна гречихи широкорядные посевы 0,60 м уступают таковым с междурядьями 0,45 м, можно предположить, что солнечная радиация и питательные вещества в разреженных посевах используются на создание вегетативной массы, а не зерна.
Таким образом, создавать оптимальную площадь листьев гречихи можно различными технологическими приёмами. Подбор лучших агротехнических условий, обеспечивающих оптимальное развитие листовой поверхности, имеет важное практическое значение.
Главным показателем фотосинтетической деятельности растений является чистая продуктивность фотосинтеза. Связь её с урожаем самая непосредственная и при равенстве площади листьев урожай сухой биомассы растений прямо пропорционален чистой продуктивности фотосинтеза (Т.М. Русакова, 1974; Schilling Herbert, 1980; J.H. Golbeck, 1992).
Продуктивность фотосинтеза гречихи не остается постоянной в течение всего периода вегетации. Наибольшую величину она имеет в период бутонизации-цветения, то есть в период наибольшего прироста сухой массы растений (Т.М. Русакова, 1974). Чистая продуктивность фотосинтеза следует за уровнем минерального питания: наиболее высокие показатели её наблюдаются у растений на чернозёмной почве с внесением минеральных удобрений (7,44 г/м2 в сут. в среднем за вегетацию).
Внесение минеральных удобрений в опытах Т.М. Русаковой (1974) довольно результативно отразилось на величине чистой продуктивности фотосинтеза. Это можно объяснить тем, что хорошее минеральное и, прежде всего, азотное питание активизирует фотосинтез деятельных, хорошо освещённых листьев верхних и средних ярусов, которые в сильной степени поддерживают активную жизнедеятельность листьев нижних ярусов. Последние, в условиях ослабленного света и затруднённого минерального питания, обычно быстро стареют и отмирают. Очевидно, поэтому на высоких фонах минерального питания образуются растения с большой площадью листьев и с высокими показателями чистой продуктивности фотосинтеза.
Анализ результатов определения чистой продуктивности фотосинтеза гречихи в условиях Новосибирской области говорит о том, что изменение этого показателя во времени и по способам возделывания культуры происходит по аналогии с изменением площади листьев и фотосинтетического потенциала. При этом, независимо от способа возделывания, чистая продуктивность фотосинтеза всегда была выше в благоприятные по увлажнению годы (А.Д. Гончаров, 2008).
По мнению Н.М. Городнего (1980), гречиха имеет лучшую, чем многие культуры способность к синтезу органических кислот (в мг на 1 г сухого вещества растения): гречиха – 7,01; нут – 5,08; люпин синий – 4,60; горчица – 4,55; горох – 3,02; овес – 2,88; кукуруза – 1,38. Всё это позволяет считать гречиху нетребовательной к почвам и приводит к тому, что её часто высевают на малоплодородной почве, где другие культуры почти не дают урожая.
По подсчетам А.А. Ничипоровича (1959, 1966), посевы культурных растений в процессе фотосинтеза способны образовывать до 8–10 г сухой биомассы на 1 м2 листовой поверхности, а потенциальная продуктивность может достигать 20–40 г/м2
в сутки. Этот показатель зависит от освещённости солнцем, притока питательных веществ и воды.
Гречиха имеет много точек роста, которые потребляют большое количество продуктов ассимиляции. Ассимиляция проходит при фотосинтезе, который напрямую зависит от величины листовой поверхности и интенсивности солнечного освещения. Величина листовой поверхности гречихи по отношению к количеству цветков на растении составляет всего 0,30–0,56 см2 на один цветок. Эта площадь слишком мала для полного обеспечения цветка пластическими веществами. Если период быстрого роста побегов совпадает со временем развития цветков и плодов, то при недостатке продуктов ассимиляции растущие верхушки стеблей и ветвей оказываются обеспеченными пищей лучше, чем большая часть репродуктивных органов (Н.М. Городний, 1980; J. Oppereer, 1985; S. Wojcik, 1991).
Листья гречихи не обладают высокой энергией усвоения углекислоты. При высокой температуре днём, даже при достаточной влагообеспеченности растений, листья подвядают. Фотосинтез ухудшается из-за быстрого роста вегетативных органов. В результате листья нижнего яруса сильно затеняются и освещаются рассеянным светом, существенно потерявшим физиологически активные лучи. Это снижает образование пластических питательных веществ, недостаток которых вызывает отмирание цветков и завязей (Vong Hguyen Quoc, 1977; И.Н. Елагин, 1984).
Гречиха – светолюбивое растение. Чем лучше освещены её листья, тем энергичнее и больше она усваивает из воздуха углекислоту, перерабатывает её в легкоусвояемые питательные вещества.
В условиях почвенной засухи рост гречихи прекращается, но развитие её продолжается, причем происходит оно быстрее, чем в нормальных условиях. В результате образуются карликовые растения, которые быстро отцветают и созревают, а урожай зерна резко снижается (Н.М. Городний, 1980).
По мнению М.Н. Городнего (1980), на загущенных посевах верхний ярус листьев гречихи поглощает только 60–70 % солнечной радиации. Средний и нижний ярусы получают третью часть и меньше общей энергии солнечного света. В среднем ярусе широкорядного посева проникающая радиация составляет 60–80 % общей падающей, а в нижнем – 50–70 %. Разреженный посев лучше использует световой режим для процессов фотосинтеза, по сравнению с загущенным.
Известно, что при затенении нижних ярусов листьев растений чистая продуктивность фотосинтеза начинает уменьшаться. Лучший по количеству и качеству урожай зерна гречихи можно получить только в посевах, обладающих достаточной по размерам площадью листьев – 40–45 тыс. м2/га при оптимальном ходе её наращивания (М.Н. Городний, 1980). В наших исследованиях лучшей ассимилирующей поверхностью гречихи был показатель на уровне 56,7 тыс. м2/га. Расчёты говорят о том, что в процессе фотосинтеза гречиха может формировать за сутки до 7,34 г сухой биомассы на 1 м2 листовой поверхности. При этом лучшие показатели урожайности отмечались при среднесуточной продуктивности фотосинтеза – 5,83 г сухой биомассы на 1 м2.
Таким образом, сравнение показателей ассимиляционной поверхности и фотосинтетической деятельности растений гречихи между вариантами, позволяет сделать вывод о том, что способы посева и нормы высева оказывают определяющее влияние на урожай зерна. Эти агротехнические приёмы можно отнести к наиболее результативным.
Фотосинтетический потенциал посевов гречихи является обобщающим показателем воздействия многих агротехнических факторов, в том числе способа посева и нормы высева. Н.Д. Кумскова (2004) считает, что фотосинтетический потенциал гречихи может составлять от 1,076 до 1,439 млн. м2 дней /га в зависимости от вносимых удобрений.
По мнению Н.М. Городнего (1980), основная причина отмирания большей части репродуктивных органов гречихи заключается в слабом снабжении их пластическими веществами. Невысокая озернённость гречихи обусловлена неудовлетворительным питанием её цветков из-за низкой облиственности растения и одновременного развития вегетативных и генеративных частей. Продукты синтеза перемещаются в растениях к тем органам, где они потребляются, и в том количестве, в котором они используются. Чем активнее жизнедеятельность организма и чем он мощнее, тем большее количество продуктов ассимиляции поступает в растение. Если образуется много растущих органов и общая потребность их в органическом веществе превысит синтетическую деятельность ассимиляционного аппарата растения, у такого растения наблюдается недостаток продуктов ассимиляции и его растущие части голодают. При этом больше страдают органы, которые растут менее интенсивно, в которые органические вещества поступают слабее. Их рост приостанавливается, но если недостаток питания длителен или даже постоянен, растения гибнут.
Бороздковые посевы гречихи в условиях Новосибирской области, в сравнении со сплошными рядовыми, формировали более значительный фотосинтетический потенциал. При этом преимущество бороздковых посевов по данному показателю последовательно нарастало от ранних периодов роста и развития растений, к более поздним. В среднем за 3 года исследований А.Д. Гончарова (2008), за период всходы-бутонизация, фотосинтетический потенциал бороздковых посевов был больше соответствующего показателя сплошных рядовых на 15 %, а за период бутонизация-цветение – на 28 %, за период цветение-созревание – на 30 %.
В опытах Н.Д. Кумсковой (2004) высокая облиственность растений и урожайность гречихи отмечены на посевах нормой 3 млн. всх. зёрен на 1 га. При высеве 2 млн. семян снижался выход зерна из-за недостаточной густоты стояния растений, что приводило к неэффективному использованию пашни. Нормы высева 4 и 5 млн. зёрен нерациональны, так как в таких посевах формировалась избыточная вегетативная масса. Наиболее высокая масса листьев гречихи, по отношению к массе растений (облиственность), наблюдалась в фазу бутонизации. Затем облиственность растений гречихи к началу цветения снижалась в два раза, достигала минимального значения в фазу созревания. На делянках, удобренных азотом, снижение её было более интенсивным, чем на контроле и других фонах. В этом варианте облиственность к моменту созревания равнялась 13,3 %, вместо 18,6 % на контроле.
Полученные нами данные для лесостепной зоны Алтайского края не противоречат результатам исследований Н.Д. Кумсковой (2004).
Наши наблюдения говорят о том, что максимального значения фотосинтетический потенциал достигал в разреженном травостое в фазу плодообразования, то есть при широкорядном способе посева (табл. 59).
Фотосинтетический потенциал и облиственность гречихи посевной
(средние за 2010–2011 гг.)
Читайте также: