
Для идентификации сортов пшеницы используют глиадины
Обновлено: 07.09.2024
Ключевые слова
запасные белки семян; электрофорез белков; международные стандартные лабораторные методы семенного контроля
Полный текст:
Литература
Идентификация сортов и регистрация генофонда культурных растений по белкам семян / Под ред. В. Г. Конарева. — СПб.: ВИР, 2000. С. 3 – 181.
Конарев В. Г. Белки растений как генетические маркеры. — М.: Колос, 1983. С. 5 – 25.
Конарев В. Г. / Тр. по прикл. бот., генет. и селек. 1987. Т. 114. С. 3 – 14.
Конарев В. Г. Морфогенез и молекулярно-биологический анализ растений. — СПб.: ВИР, 2001. С. 172 – 195.
Конарев А. В. / Аграрная Россия. 2006. № 6. С. 4 – 22.
Конарев В. Г., Гаврилюк И. П., Губарева Н. К. Способ сортовой идентификации зерна и муки. А. с. № 507271. Заявка 1 сентября 1972 г.; опубл. 11 ноября 1975 г. Бюл. № 11.
Cook R. J. The standartizations of electrophoresis methods for variety identification / Biochemical Identification of Varieties: Mater. of 3rd Int. Symp. ISTA, USSR. — Leningrad: Izd. VIR, 1988. P. 14 – 27.
Konarev V. G., Gavriljuk I. P. (eds.). Biochemical Identification of Varieties: Mater. of 3rd Int. Symp. ISTA., 1987. — Leningrad, 1988. P. 28 – 257.
Int. Rules for Seed Testing. Rules 1996. Verification of species and cultivar / Seed Sci. Technol. 1996. Suppl. 24. P. 253 – 270.
Рекомендации по использованию белковых маркеров в сортоиспытании, семеноводстве и семенном контроле / Под ред. В. Г. Конарева. — М. – Л.: Госагропром СССР, ВИР, 1989. С. 2 – 18.
Созинов А. А. Полиморфизм белков и его значение в генетике и селекции. — М.: Наука, 1985. С. 202 – 223.
Конарев В. Г., Гаврилюк И. П., Губарева Н. К. Сортовая идентификация и регистрация генетических ресурсов пшеницы по электрофоретическому спектру глиадина / Генетические ресурсы пшеницы. — Л., 1976. С. 113 – 120.
Woychik J. H., Boyndy J. A., Dimler R. J. Starch-gel electrophoresis of wheat gluten proteins with concentrated urea / Arch. Biochem. Biophys. 1961. V. 94. P. 477 – 482.
Конарев А. В., Семихов В. Ф., Примак С. П., Арефьева Л. П. О составе спирторастворимой фракции белков семян злаков / С.-х. биол. 1984. № 7. С. 13 – 17.
Губарева Н. К., Юмагузина Х. А., Павлова Н. Е. / Тр. по прикл. бот., генет. и селек. 1987. Т. 114. С. 76 – 81.
Зеленская Я. Г., Конарев А. В., Лоскутов И. Г. и др. / Аграрная Россия. 2004. № 6. С. 50 – 58.
Конарев В. Г., Губарева Н. К., Гаврилюк И. П. / Бюл. ВИР. 1982. Вып. 119. С. 60 – 63.
Губарева Н. К., Руденко М. И., Чернобурова А. Д. / Тр. по прикл. бот., генет. и селек. 1979. Т. 63. Вып. 3. С. 24 – 31.
Конарев А. В., Губарева Н. К., Корнюхин Д. Л., Бернер А. / Аграрная Россия. 2004. № 6. С. 30 – 33.
Губарева Н. К., Мартыненко Н. М., Зуев Е. В., Брыкова А. В. / Тр. по прикл. бот., генет. и селек. 2012. Т. 170. С. 158 – 162.
Пюккенен В. П., Губарева Н. К., Митрофанова О. П. / Аграрная Россия. 2005. № 2. С. 31 – 35.
Алпатьева Н. В., Губарева Н. К. / Аграрная Россия. 2002. № 3. С. 28 – 30.
Алпатьева Н. В., Губарева Н. К. / Аграрная Россия. 2002. № 3. С. 24 – 27.
Laemmli U. K. Cleavage of structural protein during assembly of the head of bacteriophage / Natura. 1970. V. 227. No. 4. P. 680 – 685.
Применение электрофореза белков в первичном семеноводстве зерновых культур: метод. указания / Под ред. В. Г. Конарева и В. Г. Еникеева. — СПб.: ВИР, 1993. С. 18 – 25.
Casarini E., Vicario A. L. / ISTA News Bul. April 2008. No. 135. P. 26 – 27.
Ссылки

В эндоспермах зерен 19 сортов мягкой пшеницы, относящихся к виду T.aestivum L. были рассмотрены идентификация глиадинкодирование аллельных локусов запасных белков и электрофоретические спектры глиадина (объем, высота, площадь и молекулярная масса). Вместе с тем был подсчитан индекс генетической схожести этих сортов по глиадин электрофоретическому спектру. После определения при помощи мономерно белковых маркеров индекса генетической схожести сортов и, проанализировав по методу SPSS статистической программы генетическую близость сортов, была составлена дендрограмма. Ключевые слова: мягкой пшеницы, глиадин, генетической схожести, глиадиновые маркеры.

Введение
Оценка селекционного материала на основе белкового маркера дает возможность достаточно быстро и качественно проводить отбор и контролировать передачу желаемых признаков от родительских форм в гибридные популяции. Вместе с тем, для селекции необходимо выявление новых и стабильных белковых маркеров для определения качества зерна пшеницы. Наиболее широко изучаемыми белками пшеницы являются запасные белки глиадин и глютенин. Полная характеристика отдельных фракций и компонентов белков отражается во многих научных исследованиях. Поэтому изучение генетической закономерности накопления белков в зерне различных сортов пшеницы и использование при идентификации генотипов пшеницы в роли маркера компонент глиадиновых белков является наиболе актуальной проблемой современности [1].
Практически ряд молекулярных маркеров используется при исследовании пшеницы. Вместе c тем, среди изученных молекулярных маркеров более досконально изучались микросателлитные SSR (Simple Sequence Repeat) маркеры ДНК.
Помимо того, что маркеры должны обладать особой спецификой, они должны быть еще и хорошо различимыми белками. Поэтому заранее изучаются биологическая специфика, молекулярное строение и другие биохимические структуры данных белков. Так как гены располагаются в различных локусах хромосом и в целом составляют геном, основываясь на принцип ген-белок, используя их как маркер, можно получить информацию о генотипе [2,14].
Хорошо известна значимость белков глиадина и глютенина в хлебопечении [12]. Качество хлеба напрямую связано с наличием или отсутствием специальных белковых единиц [8]. Кроме того, качество хлеба зависит от соотношения мономерных белков к полимерным белкам, а также от качества и объема распространения полимерных белков [9]. Глютеновые белки, глиадины и глютенины составляют 80-85% от общих белков муки и тем самым они придают эластичность и растяжимость пшеничной муке [15].
Одним из актуальнейших вопрос целевой науки является изучение синтеза генетического надзора над запасными белками глиадин и глютенин, составляющими основу эндосперма. Так как количество клейковины в зерне составляет 80-85% от запасных белков, эти белки называют белками клейковины [3,7].
Материал и методика
В качестве материала исследований использовались местные сорта мягкой пшеницы Азербайджанской Республики Акинчи-84, Гобустан, Нурлу-99, Азаметли 95, Рузи-84, Гырмызыгюль, Баяз, Парзиван-1, Парзиван-2, Шеки-1, Шафаг, Дурдана, Бирлик, Достлуг, Бол-Бугда, интродуцированные Дагдаш, Анза, Безостая-1, Саратовская-29. В зерне эндосперма этих растений были изучены запасные белки и составлены электрофореограммы глиадинкодирующих локусов Gld 1A, Gld 1B, Gld 1D, Gld 6A, Gld 6B, Gld 6D. Электрофоретический анализ проводился по методу W.Bushuk, R.R.Zillman, модифицированный Ф.А.Попереля и др. [4,5].
Результаты и обсуждения
В проводимых нами научных исследованиях наличие в генах глиадинкодирующих локусов большого количества аллелей привело к расширению каталога компонентов блоков глиадин аллелей.
Идентификация исследуемых сортов согласно генетической формуле электрофореограмм была претворена в жизнь на основе глиадинкодирующих локусов Gld 1A, Gld 1B, Gld 1D, Gld 6A, Gld 6B, Gld 6D и Gld 2-1А.
Во время проведения исследований запасные белки глиадина были условно разделены на 4 зоны и получили название ω-, γ-, β-, α-глиадины. Глиадины, находящиеся в зерне пшеницы-гетерогенные белки. Глиадины, будучи в форме мономерной цепи, составляют 50% зерна и по молекулярному весу в полиакриламидном геле имеет 4 различные формы (α-глиадин, β-глиадин, γ-глиадин, ω-глиадин) [16]. Все глиадины считаются мономерами, несмотря на то, что у них могут быть дисульфидные связи (ω-глиадин), а также цепочковидные дисульфидные связи (α-, β-, γ-глиадины) [10,11].
Была проведена идентификация компонентов блоков глиадиналлелей двух местных сортов: Экинчи-84, Эземетли-95 на основе электрофореограмм глиадинкодирующих локусов (рис. 1).
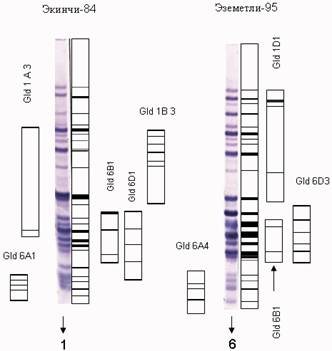
Рис. 1. Блоки компонентов аллелей в сортах мягкой пшеницы
Была определена частота встречаемости блоков компонентов аллелей в местных сортах пшеницы, таких как Акинчи-84, Gld 1A3, Gld 1B3, Gld 1D1, Gld 6A1, Gld 6B1, Gld 6D1 и Gld 2-1А3; блоков компонентов аллелей Гобустан, Gld 1A5, Gld 1B? новый, Gld 1D1, Gld 6A4, Gld 6B2, Gld 6D3 и Gld 2-1А3; Ниглу-99, Gld 1A5, Gld 1B3, Gld 1D1, Gld 6A4, Gld 6B2, Gld 6D2 и Gld 2-1А3; а также в сорте Гырмызыгюль, Gld 1A5, Gld 1B1, Gld 1D1, Gld 6A2, Gld 6B2, Gld 6D2. Однако, в сорте Гырмызыгюль Gld 2-1А3 блоков компонентов аллелей не встречалось.
Во время проведения электрофоретического анализа стало известно, что в зернах сортов мягкой пшеницы Гобустан, Нурлу-99, Рузи-84, Гырмызыгюль, Дурдана были идентифицированы глиадинкодирующие локусы, отвечающие за синтез запасных белков Gld 1A5, в сорте Азаметли-95, Gld 1A6, в сортах Парзиван-1, Шеки-1, Достлуг, Бирлик, Дагдаш, Безостая-1 (St) и Анза Gld A4; в сортах Акинчи-84, Бол-бугда, Баяз Gld 1A10, а в сорте Шафаг Gld 1A2. Таким образом, в местных сортах мягкой пшеницы частота повторяемости блоков компонентов глиадин аллелей различна. В этих сортах блоки компонентов аллелей Gld 1A5 по сравнению с блоками Gld 1A4 и Gld 1A6 встречаются довольно чаще. Блок Gld 1A5 в азербайджанских сортах встречается чаще, поэтому высокий показатель качества семян этих сортов соответственно выше. Это дает основание сделать вывод, что качество этих семян есть молекулярный белковый маркер.
В результате вышеизложенного можно заключить, что частота встречаемости глиадинкодирующих локусов аллелей в мягкой пшенице связана с генетическим аппаратом, этих же глиадинкодирующих локусов.
В вышеуказанных сортах Экинчи-84, Нурлу-99, Эземетли-95 и Дурдана Gld 1B3, Гырмызыгул, Рузи-84, Безостая-1, Анза, Бирлик, Достлуг, Перзиван-2, Шефег, Дагдаш Gld 1B1 была проведена идентификация блоков компонентов. В сортах мягкой пшеницы Экинчи-84, Саратовская-29, Бол-бугда, Беяз, парзиван-1 Gld 6D1, Нурлу-99, Гырмызыгуль, Анза, Дурдана Gld 6D2, Гобустан, Эземетли-95, Рузи-84, Дагдаш, Бирлик, Достлуг, Перзиван-2, Шеки-1 Gld 6D3, Безостая-1 (St), Шефег Gld 6D4 была проведена идентификация блоков компонентов аллелей.
Gld 1A4 блок компонентов наблюдался у классических сортов Безостая-1 (St), Дагдаш (Турция), Парзиван-2, Шеки-1. На основании литературных данных этот блок является маркерам качестве хлеба [6].
Наряду с хромосомой Gld 1A глиадинкодирующих локусов выявлен локус Gld 2-1A, который локализуется там же и контролирует синтез этого спектра.
В сортах Экинчи-84, Гобустан, Нурлу-99, Эземетли-95, Рузи-84, Бол-бугда, Бейаз, Парзиван-1 Gld 2-1A1 были выявлены блоки компонентов аллелей (табл. 1).
Во время исследований местных сортов (Гобустан, Бол-бугда, Беяз и Шеки-1) были обнаружены новые блоки компонентов аллелей (рис. 2, 3).
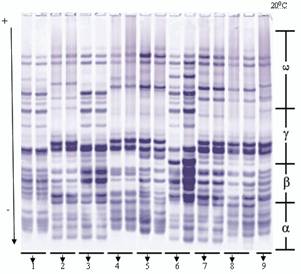
Рис. 2. Электрофореграммы глиадинокодирующих локусов мягкой пшеницы
1 - Экинчи-84, 2 - Гобустан, 3 - Нурлу-99, 4 - Гырмызыгул, 5 - Безостая (st),
6 - Эземетли- 95, 7- Анза, 8 - Рузи- 84, 9 - Саратовская-29
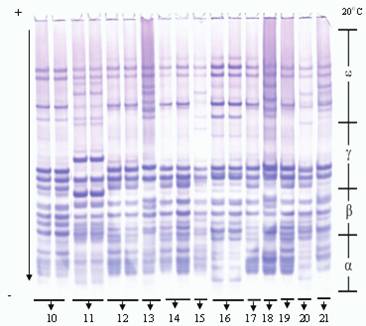
Рис. 3. Электрофореграммы запасных белков глиадина в сортах мягкой пшеницы
10 - Дагдаш, 11 - Дурдана, 12 - Бирлик, 13 - Бол-бугда, 14 - Безостая (st), 15 - Бейаз,
16 - Анза (marker), 17 - Достлуг, 18 - Парзиван-1, 19 - Парзиван-2, 20 - Шеки-1, 21 - Шефег

ИЗУЧЕНИЕ ГЕНЕТИЧЕСКОГО РАЗНООБРАЗИЯ СОРТОВ МЯГКОЙ ОЗИМОЙ ПШЕНИЦЫ ПО ГЛИАДИНКОДИРУЮЩИМ ЛОКУСАМ
© 2015 г. А. Ю. Новосельская-Драгович1, Л. А. Беспалова2, А. А. Шишкина1, В. А. Мельник1, В. П. Упелниек1, А. В. Фисенко1, Л. В. Дедова1, А. М. Кудрявцев1
Институт общей генетики им. Н.И. Вавилова Российской академии наук, Москва 119991
2Краснодарский научно-исследовательский институт сельского хозяйства им. П.П. Лукьяненко Российской академии сельскохозяйственных наук, Краснодар 350012 Поступила в редакцию 24.07.2014 г.
Методом электрофореза в ПААГе изучены 150 сортов мягкой озимой пшеницы российской и зарубежной селекции. По шести глиадинкодирующим локусам выявлено 70 аллелей. Установлено, что 42% сортов являются гетерогенными, т.е. представленными несколькими генотипами, и 58% — гомогенными. Уникальным сочетанием аллелей глиадинкодирующих локусов характеризуются 91.3% исследованных сортов, а 8.7% сортов имели одинаковые аллели по всем глиадинкодирующим локусам и не различались. Выявлены часто и редко встречающиеся аллели, причем частые аллели составляют 18.6% всех аллелей. Установлено, что разнообразие аллелей по локусам Gli-2 (47 аллелей) почти в 2 раза выше, чем по локусам Gli-1 (23 аллеля), и определяется количеством редких аллелей. Определены новые для мягкой озимой пшеницы аллели: три аллеля по локусу Gli-A2 и два аллеля по локусу Gli-B2. Выявлена тенденция снижения уровня генетического разнообразия у современных сортов вследствие использования в селекционных программах одинаковых родительских сортов.
Мягкая пшеница (ТгШеыт агзИуыт Ь.), называемая еще хлебной, является одной из основных зерновых культур РФ и занимает большую долю в пищевом рационе населения. Научная селекция продолжает создавать все новые сорта, отличающиеся улучшенными хозяйственно-ценными характеристиками. Только в 2013 г. в Государственный реестр селекционных достижений введены 14 новых сортов озимой мягкой пшеницы.
Обилие существующих сортов создает необходимость их идентификации и паспортизации, которые позволяли бы различать сорта между собой, проводить экспертизу их аутентичности, подтверждать подлинность и оценивать чистосортность образца. Основной метод установления сортовой принадлежности семян и определения сортовой чистоты — грунтовой контроль — требует больших материальных, ресурсных и временных затрат. Для идентификации коммерческих партий сортов, разрешения арбитражных споров и т.д. необходимо иметь экспресс-систему, позволяющую быстро провести анализ и получить воспроизводимые результаты. В качестве такой экспресс-системы предложен метод электрофореза запасных белков пшеницы — глиадинов [1].
Глиадины контролируются множественными аллелями шести независимых локусов, расположенных в хромосомах первой и шестой гомеоло-гических групп геномов А, В и Э [2]. Аллели глиа-
динкодирующих локусов контролируют блоки полипептидов, которые наследуются как единый менделевский признак. Блоки компонентов, контролируемые разными аллелями одного локу-са, различаются числом и электрофоретической подвижностью полипептидов [3—8]. Используя электрофоретические спектры глиадинов, теоретически можно выделить более миллиона генотипов и провести их идентификацию, что свидетельствует о высокой эффективности метода.
Для аллелей глиадинкодирующих локусов создана специальная номенклатура и разработан каталог [8]. Согласно номенклатуре локус обозначается тремя латинскими буквами (Gli), затем прописной буквой — геном (A, B или D), далее указывается порядковый номер локуса (1 или 2) и строчной буквой латинского алфавита указывается аллель, например Gli-B1k. Записывая букву аллеля каждого локуса, следующего в порядке Gli-A1, Gli-B1, Gli-D1, Gli-A2, Gli-B2, Gli-D2, получают глиадиновую формулу сорта, которая может быть использована в качестве одной из его характеристик. Эталонным признан спектр сорта Безостая 1 (глиадиновая формула Gli-A1b, Gli-B1b, Gli-D1b, Gli-A2b, Gli-B2b, Gli-D2b, или сокращенно b.b.b.b.b.b.). Он используется для определения аллелей глиадинкодирующих локусов у других сортов. Множественные алллели одного локуса записываются через знак "плюс", например Gli-D1(b + g), что
означает, что в локусе Gli-D1 присутствуют два ал-леля b и g.
Стабильность электрофоретических спектров глиадинов позволяет использовать их в качестве надежных генетических маркеров [3]. Электро-форетический спектр глиадина стабилен независимо от условий выращивания, репродукции, воспроизводим для каждого конкретного образца, сорта, партии [6, 9], являясь своего рода "отпечатком пальца" для образца. Стабильность блоков компонентов, а, следовательно, и стабильность глиадинкодирующих локусов, позволяет исследовать процессы формирования генетической изменчивости и генетического разнообразия пшеницы, оценивать генетическое разнообразие и его динамику во времени и проводить мониторинг генетической изменчивости [10—12].
Цель нашей работы — изучение генетического разнообразия 150 сортов озимой мягкой пшеницы на основе анализа электрофоретических спектров глиадина, контролируемого множественными аллелями глиадинкодирующих локусов, и определение генетической структуры сортов.
МАТЕРИАЛЫ И МЕТОДЫ
В работе были изучены 150 сортов озимой мягкой пшеницы, включенных в "Государственный реестр селекционных достижений, допущенных к использованию" (за 2012 г.). Образцы сортов были получены от оригинаторов из девяти регионов, различающихся природно-климатическими характеристиками. Из каждого региона отобрана репрезентативная выборка сортов. Для сравнения взяты сорта, созданные на Украине, но возделываемые на территории РФ и внесенные в Госреестр Российской Федерации. Одиннадцать сортов получены от оригинаторов из Западно-Сибирского региона, 35 сортов — из Зернограда, 51 — из Краснодара, 12 — от оригинаторов Нечерноземной полосы, восемь — из Саратова, 14 — из региона Средней Волги, три — из Ставрополья, шесть — из Уральского региона, три — от оригинаторов Центрально-Черноземной зоны, один — из Кургана, шесть образцов из Украины (г. Одесса) (табл. 1).
Идентификацию аллельных вариантов блоков компонентов глиадинов проводили в ПААГ (pH буфера 3.1) согласно методике, разработанной в ИОГен РАН [4, 13]. Глиадины индивидуальных зерновок экстрагировались 70%-ным этанолом. Для исследования брали не менее 100 зерновок каждого сорта. Идентификацию аллелей глиадинкодирующих локусов осуществляли в соответствии с каталогом этих аллелей [8, 13]. В качестве сортов-стандартов, относительно которых определялись аллели глиадинкодирующих локусов, использовались сорта Безостая 1 и Миронов-
ская 808. Для обозначения аллелей глиадинкодирующих локусов применяли Международную номенклатуру [14].
Всего проанализировано более 15000 электрофоретических спектров отдельных (индивидуальных) зерновок. На их основе охарактеризована генетическая структура 150 сортов озимой мягкой пшеницы.
Определение состава и генетических формул сортов
В результате анализа электрофоретических спектров глиадина был установлен аллельный состав глиадинкодирующих локусов у 150 сортов озимой мягкой пшеницы (табл. 1).
Всего по шести глиадинкодирующим локусам определено 70 аллелей. По локусу Gli-A1 идентифицировано 10 аллельных вариантов: a, Ь, c, f, g, h, I, m, o, q; по локусу Gli-B1 — шесть: Ь, d, e, f, g, I; по локусу Gli-D1 — семь: a, Ь, d, f, g,], I. По локусу Gli-A2 идентифицировано 18 аллельных вариантов блоков (а, Ь, с, d, e, f, g, i,у, k, I, m, п, p, г; new1, new2, new3), по локусу Gli-B2 — 16 (Ь, с, d, e, g, h, i, ], m, new1, o, q, t, u, v, new2), по локусу Gli-D2 — 13 аллельных вариантов (а, Ь, с, ^ e,f h,], k, m, п,p, q). Аллели new1, new2, new3 — это не идентифицированные ранее аллели, не включенные в опубликованный каталог аллелей глиадинкодирующих ло-кусов (рис. 1).
В целом аллели локусов первой гомеологиче-ской группы хромосом (ЭН-А1, Gli-B1 и Gli-D1) кодируют 23 блока компонентов, а шестой (Gli-A2, Gli-B2 и Gli-D2) группы — 47 блоков компонентов. Таким образом, локусы шестой гомеологической группы хромосом обладают большим аллельным разнообразием, они кодируют в 2 раза больше аллелей, чем локусы первой гомеологической группы. Наименьшее число аллелей характерно для локуса Gli-B1 — шесть.
Определены сорта, обладающие несколькими аллельными вариантами по одному локусу. Такие сорта называются гетерогенными, они состоят из нескольких биотипов (табл. 1). Например, сорт Августа, обладая двумя аллелями по одному локусу — Gli-D1, имеет два биотипа — Ь.Ь.].т.о.Ж и Ь.Ь^.т.о.Ж, различающихся аллелями локуса Gli-D1. Если по два аллеля имеют два локуса, то биотипов будет четыре и т.д.
Из 150 изученных сортов 63 сорта (42%) были гетерогенными по своей генетической структуре. 38 сортов состояли из двух биотипов, 17 сортов — представлены четырьмя биотипами, один сорт — шестью, пять сортов — восьмью биотипами. Сорт Альбатрос одесский содержит 24, а сорт Имени Раппопорта — 32 биотипа (табл. 1).
Таблица 1. Генетические формулы 150 гетерогенных и гомогенных сортов мягкой озимой пшеницы
Аллели глиадинкодирующих локусов
сорта ОН-А1 ОИ-Б1 он-т ОН-А2 ОП-Б2 ОН-Б2
1 Краснодар Айвина Ь 1 я п V ]
2 Краснодар Афина двуручка Ь 1 ) / Ь р
3 Зап. Сибирь Багратионовская / Ь я п т е
4 Краснодар Батько я Ь я / Ь ]
5 Урал Башкирская 10 1 Ь я т с е
6 Сред. Волга Безенчукская 380 / Ь я п т е
7 Краснодар Булгун я 1 я / т с
8 Краснодар Васса Ь 1 Ь п о Ь
9 Краснодар Веда я 1 ] / V Ь
10 Саратов Виктория 95 / Ь я п т а
11 Краснодар Вита / 1 Ь / т Ь
12 Сред. Волга Волжская С.-З. с Ь я / т а
13 Сред. Волга Волжская 100 о 1 / I пет р
14 Сред. Волга Волжская К а Ь я т Ь Ь
15 Краснодар Восторг Ь 1 Ь я Ь р
16 Нечерноз. Галина / Ь я п Ь е
17 Краснодар Гром Ь с1 я Ь Ь Ь
18 Саратов Губерния / 1 я п Ь Ь
19 Краснодар Дока т 1 а Ь е ]
20 Зерноград Дон 107 Ь Ь ] / о Ь
21 Зерноград Дон 85 / Ь я 1 т е
22 Зерноград Донская остистая Ь Ь а т Ь Ь
23 Зерноград Донская юбилейная Ь Ь я Ь Ь Ь
24 Зап. Сибирь Жатва Алта
Для дальнейшего прочтения статьи необходимо приобрести полный текст. Статьи высылаются в формате PDF на указанную при оплате почту. Время доставки составляет менее 10 минут. Стоимость одной статьи — 150 рублей.
ДЕДОВА Л.В., КУДРЯВЦЕВ А.М., МЕЛЬНИК В.А., НОВОСЕЛЬСКАЯ-ДРАГОВИЧ А.Ю., УПЕЛНИЕК В.П., ШИШКИНА А.А. — 2014 г.
КУДРЯВЦЕВ А.М., ЛЯПУНОВА О.А., МЕЛЬНИКОВА Н.В., МИТРОФАНОВА О.П. — 2010 г.
ИМАШЕВА А.Г., НОВОСЕЛЬСКАЯ-ДРАГОВИЧ А.Ю., ПУХАЛЬСКИЙ В.А., ФИСЕНКО А.В. — 2007 г.

Варочные свойства макаронных изделий зависят от содержания белка, клейковины и ее качества [1]. Качество клейковины, определяемое ее реологическими свойствами, имеет более тесную связь с варочными свойствами макарон, чем другие компоненты этой триады [2]. В свою очередь реологические свойства, тестируемые показателями SDS седиментации, миксографа, фаринографа, индексом глютена, тесно взаимосвязаны с компонентами спектра глиадинов [3], свойствами глютениновой фракции белка [4], соотношением глиадины/глютенины [5], аллельным составом глиадиновых и глютениновых локусов [6]. Глиадины и глютенины кодируются комплексными локусами Cli–1, Cli–2 (глиадины), Clu-1 (высокомолекулярные глютенины — HMW-GS) и Clu -3 (низкомолекулярные глютенины LMW-GS) тесно сцепленного с локусом Cli-1. [6]. Сорта твердой пшеницы по аллеломорфному составу локусов (Cli-B1 / CluB3) обычно распределяются на два типа. Первый тип объединяет γ — глиадин 42 (γ-42), ассоциированный с низкомолекулярным глютенином первого типа (LMW — 1). Второй тип γ — глиадин 45 (γ-45) ассоциирован с низкомолекулярным глютенином второго типа (LMW -2). Функционально качество клейковины в этих ассоциациях связано с глютениновыми компонентами, маркирующие свойства глиадиновых компонентов объясняются сцеплением с ними. Кроме того, в последнее десятилетие установлено положительное влияние субъединиц высокомолекулярных глютенинов (HMW-GS) при сочетании аллеломорфных состояний (по номенклатуреNieto-Taladrzetal, 1997) 6+8, 7+8 локуса Clu-B1 и a локуса Clu-A3 [6].
Таким образом, высокое качество клейковины формируется на широкой генетической основе. Помимо генетических факторов формирование свойств клейковины в значительной степени зависит от условий среды [7]. Очевидно, что генотипы с максимальной концентрацией положительно влияющих на качество клейковины аллелей глютениновых локусов и широким отношением глютениновой фракции белков к глиадиновой, будут стабильнее на фенотипическом уровне в разных условиях среды. Поиск таких генотипов и фонов для их идентификации — цель исследований, результаты которых представлены в данной публикации.
Читайте также: