
При скрещивании растения краснозерной пшеницы с растением белозерной пшеницы
Обновлено: 18.09.2024
копирование только того участка, который удовлетворяет заданным условиям, и только в том случае, если он присутствует в исследуемом образце. В отличие от амплификации
ДНК в живых организмах, (репликации), с помощью ПЦР амплифицируются относительно короткие участки ДНК. В обычном ПЦР-процессе длина копируемых ДНК- участков составляет не более 3000 пар оснований (3 kbp
[6]
). С помощью смеси различных полимераз, с использованием добавок и при определённых условиях длина ПЦР- фрагмента может достигать 20-40 тысяч пар нуклеотидов. Это всё равно значительно меньше длины хромосомной ДНК эукариотической клетки. Например, геном человека состоит примерно из 3 млрд пар оснований
ДНК-дактилоскопия или генетическая дактилоскопия представляет собой метод, используемый в судебно-медицинской экспертизе для идентификации лиц на основе уникальности последовательностей ДНК индивидуума. Метод был открыт в 1984 году британским генетиком Алеком Джеффризом. Рассматривая рентгеновские снимки ДНК, он обнаружил, что ДНК разных людей имеют уникальные последовательности нуклеотидов. Последовательности ДНК конкретного человека составляют его ДНК- профиль или генетический паспорт, который можно использовать для идентификации личности. Составление ДНК-профиля человека (ДНК-профилирование) не следует путать с полной рашифровкой его генома.
Секвенирование - это определение нуклеотидной последовательности избранного участка
ДНК. В результате секвенирования получают формальное описание первичной структуры линейной макромолекулы в виде последовательности мономеров в текстовом виде. В результате секвенирования перекрывающихся участков ДНК, получают последовательности участков генов, целых генов, тотальной мРНК и даже полных геномов организмов
Картирование - это создание схемы, описывающей порядок расположения генов на хромосоме. Генетические карты человека используются в медицине при диагностике ряда тяжелых наследственных заболеваний человека.
Полимеразная цепная реакция (ПЦР) – молекулярно-биологическая реакция, позволяющая быстро получить большое количество копий конкретного фрагмента ДНК. Искомый фрагмент может быть частью очень сложной смеси нуклеиновых кислот. Исходным материалом для ПЦР может быть даже единственная молекула ДНК.
Преимущества: Высокая специфичность; Высокая чувствительность; Возможность диагностики скрытых инфекций; Прямое определение наличия возбудителя; Высокая скорость получения анализа (4-5 часов)
Основы ПЦР: Использование термостабильной полимеразы. Полимераза достраивает специфические затравки (праймеры). Процесс состоит из повторяющихся температурных циклов.
Генная дактилоскопия— выявление вариаций в числе и длине тандемных повторов ДНК.
(количество повторяющихся элементов в выбранном участке генома)
Последовательности ДНК конкретного человека составляют его ДНК-профиль или генетический паспорт, который можно использовать для идентификации личности.
Составление ДНК-профиля человека (ДНК-профилирование) не следует путать с полной расшифровкой его генома.
Чем больше участков генома (или локусов) анализируется при составления ДНК-профиля, тем выше точность идентификации личности. В настоящее время число локусов для составления ДНК-профиля достигает 16 и более.
2.
Изменчивость как свойство, обеспечивающее возможность существования
живых систем в различных состояниях.
Жизнь представлена отдельными видами, являющимися совокупностями организмов, которые обладают свойствами наследственности и изменчивости. Эти свойства становятся основой эволюционного процесса. Механизмами, обусловливающими такой результат,
являются избирательная выживаемость и избирательное размножение особей, принадле- жащих к одному виду. В природных условиях особенно интенсивно размножение происходит в популяциях, которые являются минимальными самовоспроизводящимися группами особей внутри вида. Изменчивость — это способность дочерних организмов отличаться от родительских форм. Различают две основные формы изменчивости: фенотипическую (ненаследственную) и генотипическую (наследственную). Фенотипической или модификационной изменчивостью называют изменения фенотипа под действием факторов внешней среды без изменения генотипа. Так как при этом генотип не затрагивается, то эти изменения не наследуются. Развитие ряда признаков зависит и от влияния различных регуляторных систем организма, в первую очередь эндокринной. В генетической информации организма заложена способность развития определенных свойств и признаков. Эта способность реализуется лишь в конкретных условиях среды.
Генотипическая изменчивость — это изменчивость, связанная с изменением генотипа.
Понятно, что генотипическая изменчивость передается по наследству. Она подразделяется на комбинативную и мутационную. Комбинативная изменчивость связана с получением новых комбинаций имеющихся в генотипе генов. Мутационная изменчивость — это скачкообразное и устойчивое изменение генетического материала, передающееся по наследству.

6 Смотреть ответы Добавь ответ +10 баллов

Ответы 6

1 вопрос:
а) с наступлением холодной погоды давление повышается т.к воздух охлаждается и становится тяжёлым и сильнее давит на земную поверхность
б) с наступлением тёплой погоды давление понижается т.к воздух нагревается и становится лёгким и поднимается вверх оказывая не сильное давление на земную поверхность
2 вопрос:
а) на высоте 7455 м.
Нормальное давление 760 мм.рт.ст.,
(760 - 50) х 10,5 = 7455 м.
б) на высоте 315 м.
(790 - 760) х 10,5 = 315 м.
3 вопрос:
1) 3150 : 10,5 = 300 мм.рт.ст.
2) 740 - 300 = 440 мм.рт.ст.
4 вопрос:
а) А 741 Б 750 - из точки Б в точку А, разница составляет 9 мм.рт.ст.
б) А 763 Б 758 - из точки А в точку Б, разница составляет 5 мм.рт.ст.
в) А 754 Б 752 - из точки А в точку Б, разница составляет 2 мм.рт.ст.
г) А 755 Б 762 из точки Б в точку А, разница составляет 7 мм.рт.ст.
Ветер будет сильнее под буквой а)
Пояснение: ветер дует из высокого давления в низкое. Чем больше разница давления, тем сильнее ветер.
1) признак ген генотип
черная А АА, Аа
серая а аа
мохнатая В ВВ,Вв
гладкошест. в вв
Р АаВв × аавв
G АВ, Ав, аВ, ав ав
F АаВв, Аавв, ааВв, аавв

Круг - 360°, 1 час этого круга равен 15°, далее просто черти углы по кругу,
Учёба в школе - 75°
Домашняя работа - 30°
Работа с ПК - 7,5°
Просмотр телепередач - 37,5°
Кстати, ты где-то потерял/а 2 часа, либо в задаче так и есть
1. 9+5+2+3+0,5+2,5=23 ч - всего в распорядке дня.
2. 360°:23≈15,6° - 1 ч на круге
Теперь каждое значение (сон, школа и т.д.) умножаем на 15,6° и тем самым находим, сколько градусов будет на круговой диаграмме.
0.5•7.8=3.9° - работа с ПК
2.5•(7.8+15.6)°=58,5° - просмотр передач
Чтобы проверить, правильно ли вышло, мы должны сложить все значения и должно получиться 360°. У нас во втором действии приблизительно, поэтому будет не 360, а приблизительно.

У вас неправильно даны исходные данные, но я их уже исправил.
В 1-ом примере:
делятся ли 11100111000111000111111111 : 111?
Во 2-ом примере:
делятся ли 1110010100011100011111111 : 1110?
1-ый пример:
Переводим из десятичной в двоичную
11100111000111000111111111 = 60584447
111 = 7
60584447 делится на 7 (можете сами проверить на калькуляторе), а это значит что исходные двоичные числа тоже делятся.
Также, чтобы проверить делимость числа на 111, или на cемь в десятичной системе, нужно разбить число на подгруппы, состоящие из трех чисел, и просуммировать их, если полученная сумма делится на 111, то и начальное число тоже делится
Примеры:
1) Делится ли 10001100 на 111.
100 + 011 + 000 = 111.
ответ: делится
2) Делится ли 001010100111 на 111.
001 + 010 + 100 + 111 = 1110 (допишем к этому числу необходимое количество нулей, и еще раз просуммируем)
1110 = 111000
111 + 000 = 111
ответ: делится
2-ой пример:
1110010100011100011111111 = 30030079
1110 = 14
30030079 уже не делится на 14, понятно,что и исходные двоичные числа не делятся
Это можно проверить и другим Число делится на 14, только если оно делится на 7, и на 2. Проверка делимости на 7 уже известна, а для проверки делимости на 2, нужно просто проверить делится ли на него последняя цифра числа. Например, 936 делится на 2, так как 6 делится. В двоичной системе надо взять первые 4 разряда и проверить их делимость на 0010.
Пример:
0101 0110 1010 : 1110?
010 + 101 +101 + 010 = 1110
111 + 000 = 111 => Это число делится на 7, или 111 в двоичной.
Теперь проверим делимость на два. Возьмем первые 4 разряда - 1010.
1010 = 10
10 делится на 2.
ответ: раз число делится на 7, и на 2, то оно делится и на 14, то есть на 1110 .
Теперь обратимся к проблеме взаимодействия неаллельных генов. Если развитие признака контролируется более чем одной парой генов, то это означает, что он находится под полигенным контролем. Установлено несколько основных типов взаимодействия генов: комплементарность, эпистаз, полимерия и плейотропия.
Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур. Различные породы кур характеризуются разной формой гребня. Виандотты имеют низкий, правильный, покрытый сосочками гребень, известный под названием “розовидного”. Брамы и некоторые бойцовые куры обладают узким и высоким гребнем с тремя продольными возвышениями — “гороховидным”. Леггорны имеют простой или листовидный гребень, состоящий из одной вертикальной пластинки. Гибридологический анализ показал, что простой гребень ведет себя как полностью рецессивный признак по отношению к розовидному и гороховидному. Расщепление в F2 соответствует формуле 3 : 1. При скрещивании же между собой рас с розовидным и гороховидным гребнем у гибридов первого поколения развивается совершенно новая форма гребня, напоминающая половинку ядра грецкого ореха, в связи с чем гребень был назван “ореховидным”. При анализе второго поколения было установлено, что соотношение разных форм гребня в F2 соответствует формуле 9 : 3 : 3 : 1, что указывало на дигибридный характер скрещивания. Была разработана схема скрещивания, объясняющая механизм наследования этого признака.
В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
Наследование формы гребня у кур можно отнести к комплементарному взаимодействию неаллельных генов. Комплементарными, или дополнительными, считаются гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состоянии обусловливают развитие нового признака. Действие же каждого из генов в отдельности воспроизводит признак одного из родителей.

Схема, иллюстрирующая взаимодействие неаллельных генов,
определяющих форму гребня у кур
Наследование генов, определяющих форму гребня у кур, полностью укладывается в схему дигибридного скрещивания, так как они ведут себя при распределении независимо. Отличие от обычного дигибридного скрещивания проявляется только на уровне фенотипа и сводится к следующему:
- Гибриды F1 не похожи ни на одного из родителей и обладают новым признаком;
- В F2 появляются два новых фенотипических класса, которые являются результатом взаимодействия либо доминантных (ореховидный гребень), либо рецессивных (простой гребень) аллелей двух независимых генов.
Механизм комплементарного взаимодействия подробно изучен на примере наследования окраски глаз у дрозофилы. Красная окраска глаз у мух дикого типа определяется одновременным синтезом двух пигментов — бурого и ярко-красного, каждый из которых контролируется доминантным геном. Мутации, затрагивающие структуру этих генов, блокируют синтез либо того, либо другого пигмента. Так, рецессивная мутация brown (ген находится во 2-й хромосоме) блокирует синтез ярко-красного пигмента, в связи с чем у гомозигот по этой мутации бурые глаза. Рецессивная мутация scarlet (ген располагается в 3-й хромосоме) нарушает синтез бурого пигмента, и поэтому гомозиготы stst имеют ярко-красные глаза. При одновременном присутствии в генотипе обоих мутантных генов в гомозиготном состоянии не вырабатываются оба пигмента и глаза у мух белые.
В описанных примерах комплементарного взаимодействия неаллельных генов формула расщепления по фенотипу в F2 соответствует 9 : 3 : 3 : 1. Такое расщепление наблюдается в том случае, если взаимодействующие гены по отдельности имеют неодинаковое фенотипическое проявление и оно не совпадает с фенотипом гомозиготного рецессива. Если это условие не соблюдается, в F2 имеют место иные соотношения фенотипов.
Например, при скрещивании двух разновидностей фигурной тыквы со сферической формой плода гибриды первого поколения обладают новым признаком — плоскими или дисковидными плодами. При скрещивании гибридов между собой в F2 наблюдается расщепление в соотношении 9 дисковидных : 6 сферических : 1 удлиненная.

Схема наследования формы плода у тыкв
Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
Еще один пример комплементарного взаимодействия дает наследование окраски шерсти у мышей. Дикая серая окраска определяется взаимодействием двух доминантных генов. Ген А отвечает за присутствие пигмента, а ген В — за его неравномерное распределение. Если в генотипе присутствует только ген А (А-bb), то мыши равномерно окрашены в черный цвет. Если присутствует только ген В (ааВ-), то пигмент не вырабатывается и мыши оказываются неокрашенными, так же как и гомозиготный рецессив ааbb. Такое действие генов приводит к тому, что в F2 расщепление по фенотипу соответствует формуле 9 : 3 : 4.
Комплементарное взаимодействие описано также при наследовании окраски цветов у душистого горошка. Большая часть сортов этого растения имеет пурпурные цветы с фиолетовыми крыльями, которые характерны для дикой сицилийской расы, но есть также сорта с белой окраской. Скрещивая растения с пурпурной окраской цветов с растениями с белыми цветами Бетсон и Пеннет установили, что пурпурная окраска цветов полностью доминирует над белой, и в F2 наблюдается соотношение 3 : 1. Но в одном случае от скрещивания двух белых растений получилось потомство, состоящее только из растений с окрашенными цветами. При самоопылении растений F1 было получено потомство, состоящее из двух фенотипических классов: с окрашенными и неокрашенными цветами в соотношении 9/16 : 7/16.
Полученные результаты объясняются комплементарным взаимодействием двух пар неаллельных генов, доминантные аллели которых (С и Р) в отдельности не способны обеспечить развитие пурпурной окраски, так же как и их рецессивные аллели (ссрр). Окраска проявляется только при наличии в генотипе обоих доминантных генов, взаимодействие которых обеспечивает синтез пигмента.
В приведенном примере формула расщепления в F2 — 9 : 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление. Например, при скрещивании двух сортов кукурузы с фиолетовой окраской зерновок в F1 все гибриды имеют желтые зерновки, а в F2 наблюдается расщепление 9/16 желт. : 7/16 фиол.
Эпистаз — другой тип неаллельного взаимодействия, при котором происходит подавление действия одного гена другим неаллельным ему геном. Ген, который препятствует проявлению другого гена, называется эпистатичным, или супрессором, а тот, чье действие подавляется, гипостатичным. В качестве эпистатичного гена может выступать как доминантный, так и рецессивный ген (соответственно доминантный и рецессивный эпистаз).
Примером доминантного эпистаза служит наследование окраски шерсти у лошадей и окраски плодов у тыквы. Схема наследования этих двух признаков абсолютно одинаковая.
Схема наследования окраски шерсти у лошадей
F2
| CB | Cb | cB | cb | |
| CB | CCBB сер. | CCBB сер. | CcBB сер. | CcBb сер. |
| Cb | CCBb сер. | CCbb сер. | CcBb сер. | Ccbb сер. |
| cB | CcBB сер. | CcBb сер. | ccBB черн. | ccBb черн. |
| cb | CcBb сер. | Ccbb сер. | ccBb черн. | ccbb рыж. |
Из схемы видно, что доминантный ген серой окраски С является эпистатичным по отношению к доминантному гену В, который обусловливает черную окраску. В присутствии гена С ген В своего действия не проявляет, и поэтому гибриды F1 несут признак, определяемый эпистатичным геном. В F2 класс с обоими доминантными генами сливается по фенотипу (серая окраска) с классом, у которого представлен только эпистатичный ген (12/16). Черная окраска проявляется у 3/16 гибридных потомков, в генотипе которых отсутствует эпистатичный ген. В случае гомозиготного рецессива отсутствие гена-супрессора позволяет проявиться рецессивному гену с, который вызывает развитие рыжей окраски.

Схема наследования окраски оперения у кур
Доминантный эпистаз описан также при наследовании окраски пера у кур. Белый цвет оперенья у кур породы леггорнов доминирует над окрашенным черных, рябых и других цветных пород. Однако белая окраска других пород (например, плимутроков) рецессивна по отношению к цветному оперению. Скрещивания между особями с доминантной белой окраской и особями с рецессивной белой окраской в F1 дают белое потомство. В F2 наблюдается расщепление в соотношении 13 : 3.
Анализ схемы показывает, что в определении окраски пера у кур принимают участие две пары неаллельных генов. Доминантный ген одной пары (I) является эпистатичным по отношению к доминантному гену другой пары, вызывающему развитие окраски (C). В связи с этим окрашенное оперение имеют только те особи, в генотипе которых присутствует ген С, но отсутствует эпистатичный ген I. У рецессивных гомозигот ссii отсутствует эпистатичный ген, но у них нет гена, который обеспечивает выработку пигмента (C), поэтому они имеют белую окраску.
В качестве примера рецессивного эпистаза можно рассмотреть ситуацию с геном альбинизма у животных (см. выше схему наследования окраски шерсти у мышей). Присутствие в генотипе двух аллелей гена альбинизма (аа) не дает возможности проявиться доминантному гену окраски (B) — генотипы ааВ-.
Полимерный тип взаимодействия был впервые установлен Г. Нильсеном-Эле при изучении наследования окраски зерна у пшеницы. При скрещивании краснозерного сорта пшеницы с белозерным в первом поколении гибриды были окрашенными, но окраска была розовой. Во втором поколении только 1/16 часть потомства имела красную окраску зерна и 1/16 — белую, у остальных окраска была промежуточной с разной степенью выраженности признака (от бледно-розовой до темно-розовой). Анализ расщепления в F2 показал, что в определении окраски зерна участвуют две пары неаллельных генов, действие которых суммируется. Степень выраженности красной окраски зависит от количества доминантных генов в генотипе.
Полимерные гены принято обозначать одинаковыми буквами с добавлением индексов, в соответствии с числом неаллельных генов.
Действие доминантных генов в данном скрещивании является аддитивным, так как добавление любого из них усиливает развитие признака.
Описанный тип полимерии, при котором степень развития признака зависит от дозы доминантного гена, называется кумулятивным. Такой характер наследования обычен для количественных признаков, к которым следует отнести и окраску, т.к. ее интенсивность обусловлена количеством вырабатываемого пигмента. Если не учитывать степень выраженности окраски, то соотношение окрашенных и неокрашенных растений в F2 соответствует формуле 15 : 1.
Однако в некоторых случаях полимерия не сопровождается кумулятивным эффектом. В качестве примера можно привести наследование формы семян у пастушьей сумки. Скрещивание двух рас, одна из которых имеет треугольные плоды, а другая яйцевидные дает в первом поколении гибриды с треугольной формой плода, а во втором поколении наблюдается расщепление по этим двум признакам в соотношении 15 треуг. : 1 яйцев.
Данный случай наследования отличается от предыдущего только на фенотипическом уровне: отсутствие кумулятивного эффекта при увеличении дозы доминантных генов обусловливает одинаковую выраженность признака (треугольная форма плода) независимо от их количества в генотипе.
К взаимодействию неаллельных генов относят также явление плейотропии — множественного действия гена, влияния его на развитие нескольких признаков. Плейотропное действие генов является результатом серьезного нарушения обмена веществ, обусловленного мутантной структурой данного гена.
Так, например, ирландские коровы породы декстер отличаются от близкой по происхождению породы керри укороченными ногами и головой, но одновременно лучшими мясными качествами и способностью к откорму. При скрещивании коров и быков породы декстер 25% телят имеют признаки породы керри, 50% сходны с породой декстер, а в остальных 25% случаев наблюдаются выкидыши уродливых бульдогообразных телят. Генетический анализ позволил установить, что причиной гибели части потомства является переход в гомозиготное состояние доминантной мутации, вызывающей недоразвитие гипофиза. В гетерозиготе этот ген приводит к появлению доминантных признаков коротконогости, короткоголовости и повышенной способности к отложению жира. В гомозиготе этот ген имеет летальный эффект, т.е. в отношении гибели потомства он ведет себя как рецессивный ген.
Летальный эффект при переходе в гомозиготное состояние характерен для многих плейотропных мутаций. Так, у лисиц доминантные гены, контролирующие платиновую и беломордую окраски меха, не оказывающие летального действия в гетерозиготе, вызывают гибель гомозиготных зародышей на ранней стадии развития. Аналогичная ситуация имеет место при наследовании серой окраски шерсти у овец породы ширази и недоразвития чешуи у зеркального карпа. Летальный эффект мутаций приводит к тому, что животные этих пород могут быть только гетерозиготными и при внутрипородных скрещиваниях дают расщепление в соотношении 2 мутанта : 1 норма.
Схема наследования платиновой окраски у лис
F1
| A | a | |
| A | AA погибают | Aa платин. |
| a | Aa платин. | aa черн. |
Однако большинство летальных генов рецессивны, и гетерозиготные по ним особи имеют нормальный фенотип. О наличии у родителей таких генов можно судить по появлению в потомстве гомозиготных по ним уродов, абортусов и мертворожденных. Чаще всего подобное наблюдается в близкородственных скрещиваниях, где родители обладают сходными генотипами, и шансы перехода вредных мутаций в гомозиготное состояние достаточно высоки.
Плейотропные гены с летальным эффектом есть у дрозофилы. Так, доминантные гены Curly — загнутые вверх крылья, Star — звездчатые глаза, Notch — зазубренный край крыла и ряд других в гомозиготном состоянии вызывают гибель мух на ранних стадиях развития.
Известная рецессивная мутация white, впервые обнаруженная и изученная Т. Морганом, также имеет плейотропный эффект. В гомозиготном состоянии этот ген блокирует синтез глазных пигментов (белые глаза), снижает жизнеспособность и плодовитость мух и видоизменяет форму семенников у самцов.
У человека примером плейотропии служит болезнь Марфана (синдром паучьих пальцев, или арахнодактилия), которая вызывается доминантным геном, вызывающим усиленный рост пальцев. Одновременно он определяет аномалии хрусталика глаза и порок сердца. Болезнь протекает на фоне повышения интеллекта, в связи с чем ее называют болезнью великих людей. Ею страдали А. Линкольн, Н. Паганини.
Плейотропный эффект гена, по всей видимости, лежит в основе коррелятивной изменчивости, при которой изменение одного признака влечет за собой изменение других.
К взаимодействию неаллельных генов следует отнести также влияние генов-модификаторов, которые ослабляют или усиливают функцию основного структурного гена, контролирующего развитие признака. У дрозофилы известны гены-модификаторы, модифицирующие процесс жилкования крыльев. Известно не менее трех генов-модификаторов, влияющих на количество красного пигмента в волосе крупного рогатого скота, в результате чего окраска шерсти у разных пород колеблется от вишневой до палевой. У человека гены-модификаторы изменяют окраску глаз, усиливая или ослабляя ее интенсивность. Их действием объясняется разная окраска глаз у одного человека.
Существование явления взаимодействия генов привело к появлению таких понятий, как “генотипическая среда” и “генный баланс”. Под генотипической средой подразумевается то окружение, в которое попадает вновь возникающая мутация, т.е. весь комплекс генов, имеющихся в данном генотипе. Понятие “генный баланс” касается соотношения и взаимодействия между собой генов, влияющих на развитие признака. Обычно гены обозначают названием признака, возникающего при мутации. На самом же деле проявление этого признака часто является результатом нарушения функции гена под влиянием других генов (супрессоров, модификаторов и др.). Чем сложнее генетический контроль признака, чем больше генов участвуют в его развитии, тем выше наследственная изменчивость, так как мутация любого гена нарушает генный баланс и приводит к изменению признака. Следовательно, для нормального развития особи необходимо не только присутствие генов в генотипе, но и осуществление всего комплекса межаллельных и неаллельных взаимодействий.
Перейти к чтению других тем книги "Генетика и селекция. Теория. Задания. Ответы":

Презентация к урокам в профильных классах по теме "Взаимодействие генов".

Взаимодействие неаллельных генов
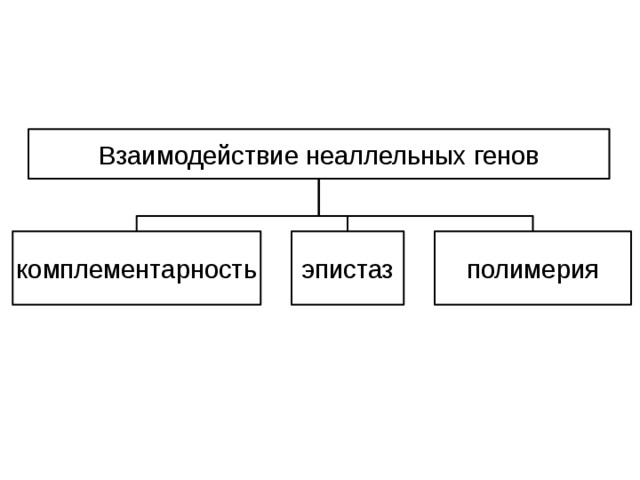
Взаимодействие неаллельных генов
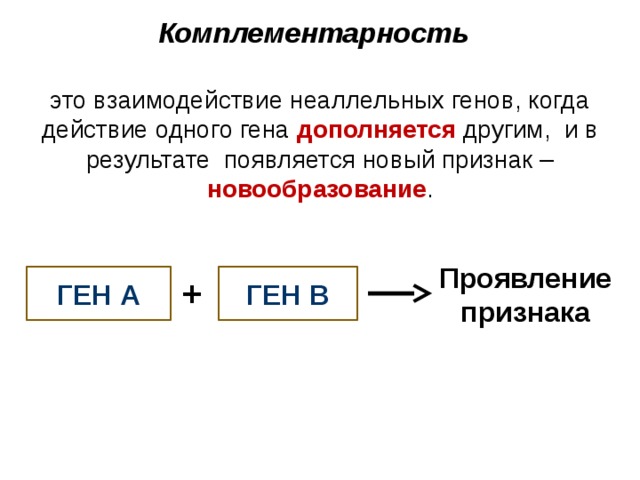
Комплементарность
это взаимодействие неаллельных генов, когда действие одного гена дополняется другим, и в результате появляется новый признак – новообразование .
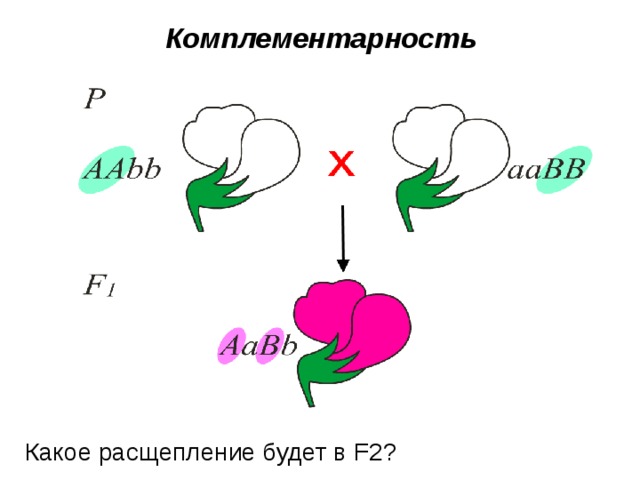
Комплементарность
Какое расщепление будет в F2?
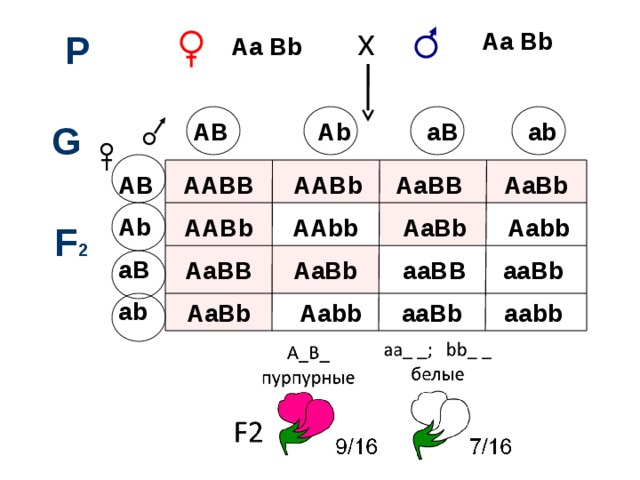
AB Ab aB ab
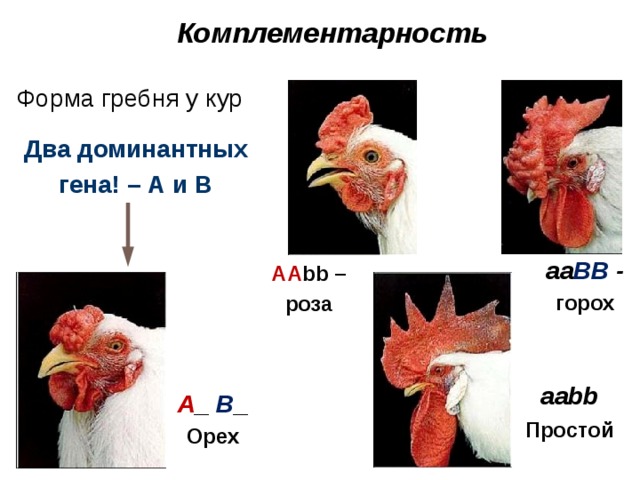
Комплементарность
Форма гребня у кур
Два доминантных гена! – А и В
aa BB - горох
АА bb –роза
A _ B _ Орех
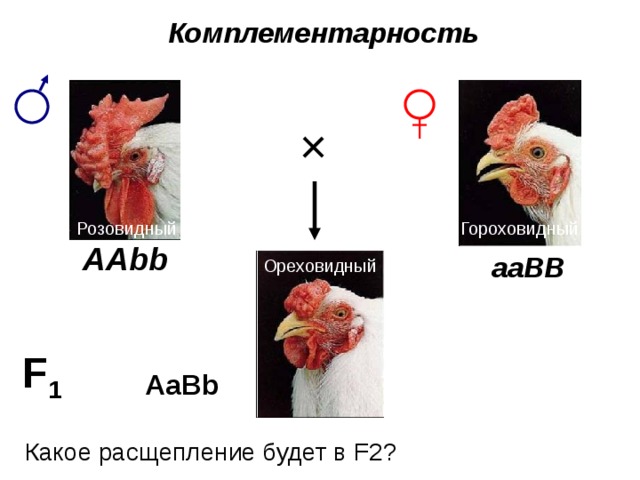
Комплементарность
Какое расщепление будет в F2?
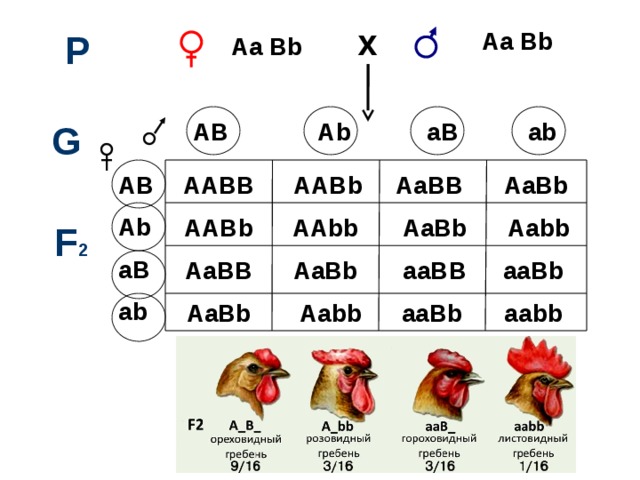
AB Ab aB ab

Эпистаз -
это взаимодействие генов, при котором
один ген подавляет действие другого неаллельного гена.
Подавляющий ген называется геном-ингибитором эпистатическим геном .
Подавляемый ген – гипостатический .
Ген А - ингибитор
Ген В - ингибитор

Эпистаз
При доминантном эпистазе расщепление по фенотипу может происходить в соотношении
При рецессивном эпистазе расщепление по фенотипу может идти в соотношении
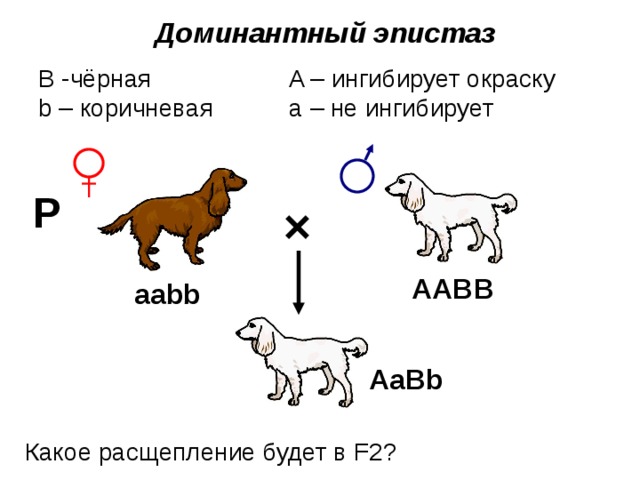
Доминантный эпистаз
B -чёрная A – ингибирует окраску
b – коричневая а – не ингибирует
Какое расщепление будет в F2?
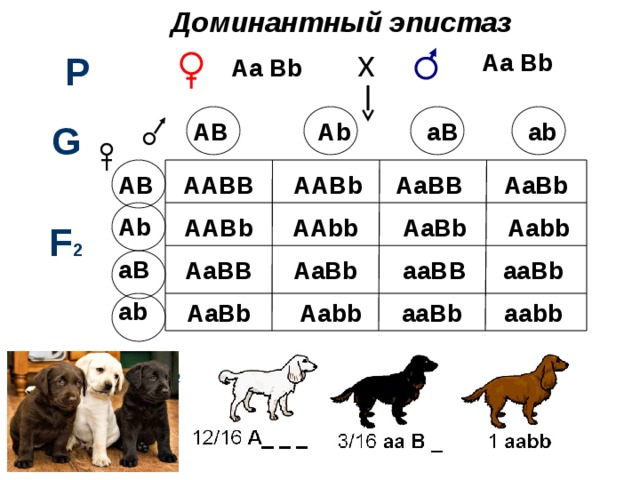
Доминантный эпистаз
AB Ab aB ab
Доминантный эпистаз
С – наличие пигмента; с – отсутствие пигмента
I – подавитель окраски; i – не подавляет
Рис. 333. Наследование окраски зерновки у пшеницы.
При скрещивании, например, леггорнов ( ССII ) с плимутроками ( ссii ), все потомство F1 имеет белую окраску, которая определяется наличием в их генотипе гена-подавителя ( СсIi ). Если же гибридов F1 скрестить между собой, то во втором поколении происходит расщепление по окраске в отношении 13/16 белых: 3/16 окрашенных. Окрашенным оказывается та часть потомства, в генотипе которой имеется ген окраски и отсутствует его подавитель ( С_ii ).
Скрещивая белую и пурпурную фасоли, Мендель столкнулся с явлением полимерии. Полимерией называют однозначное влияние двух, трех и более неаллельных генов на разви-
тие одного и того же признака. Такие гены называют полимерными , или множественными , и обозначают одной буквой с соответствующим индексом, например, А1 , А2 , а1 , а2 .
Полимерные гены контролируют большинство оличественных признаков организмов: высоту растения, массу семян, масличность семян, содержание сахара в корнеплодах сахарной свеклы, удойность коров, яйценоскость, вес тела и т.д.
Явление полимерии было открыто в 1908 г. при изучении окраски зерновки у пшеницы Нельсоном-Эле (рис. 333). Он предположил, что наследование окраски у зерновки пшеницы обусловлено двумя или тремя парами полимерных генов. При скрещивании краснозерной и белозерной пшеницы в F1 наблюдалось промежуточное наследование признака: все гибриды первого поколения имели светло-красное зерно. В F2 происходило расщепление в отношении 63 краснозерных на 1 белозерное. Причем краснозерные зерновки имели разную интенсивность окраски — от темно-красной до светло-красной. Исходя из наблюдений, Нельсоном-Эле определил, что признак окраски зерновок обуславливает три пары полимерных генов.
У человека по типу полимерии наследуется, например, окраска кожи.
Плейотропией называют множественное действие генов. Плейотропное действие генов имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет не только развитие данного признака, но и воздействует на вторичные реакции биосинтеза различных других признаков и свойств, вызывая их изменение.
Плейотропное действие генов впервые было обнаружено Г. Менделем, который обнаружил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У человека встречается рецессивная наследственная болезнь—серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов. Одновременно с этим возникают глубокие нарушения в сердечно-сосудистой, нервной, пищеварительной, выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию погибает в детстве.
Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены.
Читайте также: